 |


Отличия половых клеток от соматических
|
|
|
|
К занятию №2
ПРОГЕНЕЗ.
Началу индивидуального развития предшествует возникновение половых клеток, т.е. гаметогенез, который можно рассматривать прогенезом при индивидуальном развитии.
Процесс образования мужских половых клеток - сперматогенез. В результате образуются сперматозоиды.
Сперматозоид, спермий (spermium) состоит из головки, шейки и хвостика и содержит минимальное количество цитоплазмы, покрыт плазмалеммой.(рис. 1). В головке находится ядро с конденсированным хроматином, окруженное узким ободком цитоплазмы и клеточной оболочкой. На переднем полюсе ядра в чехлике располагается акросома – это видоизмененный комплекс Гольджи. Акросома содержит набор ферментов, среди которых основными являются протеазы и гиалуронидаза, они растворяют оболочки яйцеклетки при оплодотворении. В шейке сперматозоида располагаются проксимальная и дистальная центриоли. От дистальной центриоли начинается осевая нить хвостика, она представляет собой комплекс микротрубочек 9 пар периферических и 2 центральных.
В хвостике выделяют связующую (промежуточную), главную и конечную (терминальную) части. В связующем отделе вокруг осевой нити спирально лежат митохондрии, они обеспечивают энергетику движений сперматозоида и энергетические потребности клетки. Главная часть устроена как жгутик или ресничка и состоит из микротрубочек и опорных фибрилл. Конечный отдел хвостика единичные микротрубочки жгутика, опорные фибриллы лежат беспорядочно, а кончик хвостика образован лишь плазмалеммой и цитоплазмой.

Рис.1. Строение сперматозоида.
1-головка, 2-шейка, 3-связующий отдел, 4-главная часть, 5- акросома, 6-чехлик, 7-ядро, 8-проксимальная центриоль, 9-дистальная центриоль, 10-митохондрии, 11-осевая нить
|
|
|
Сперматозоиды животных отличаются друг от друга формой головки и соотношением указанных отделов. Длина сперматозоидов у разных животных неодинакова. Так, сперматозоид морской свинки имеет длину 100 мкм, быка – 65 мкм, воробья – 200 мкм, крокодила – 20 мкм, человека – 53 мкм.

|
Рис.2. Сперматозоиды позвоночных:
1 – щуки, 2-саламандры, 3-голубя, 4-ехидны, 5- полевой мыши
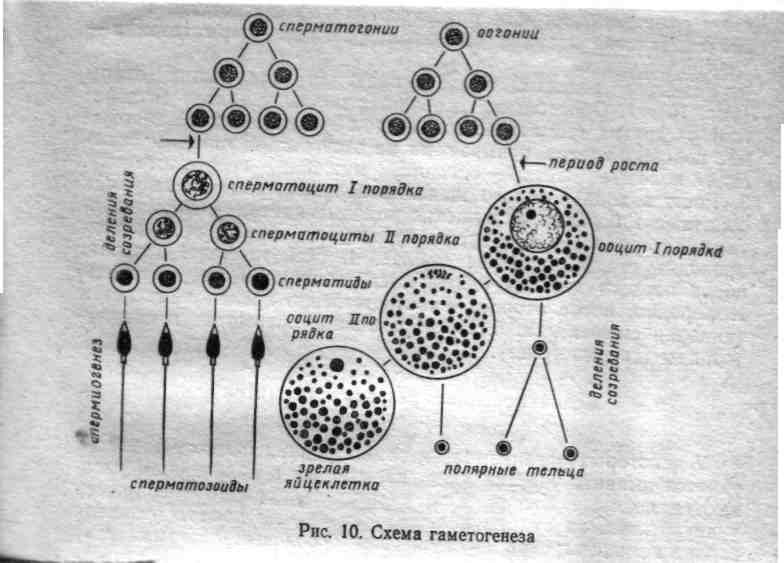
Сперматогенез
Сперматогенез происходит в извитых семенных канальцах семенников.
В сперматогенезе различают 4 периода: размножение, рост, созревание (мейоз) и формирование.
В период размножения исходные недифференцированные половые клетки - сперматогонии, или гонии делятся путем обычного митоза. Проделав несколько таких делений, они вступают в период роста. На этой стадии их называют сперматоцитами I порядка. Ониусиленно ассимилируют питательные вещества, укрупняются, претерпевают глубокую физико-химическую перестройку, в результате которой подготавливаются к третьему периоду - созреванию, или мейозу. Биологический смысл мейоза – это образование половых клеток с гаплодным набором хромосом.
В мейозе сперматоциты I проходят два процесса клеточного деления. В первом делении (редукционном) происходит уменьшение числа хромосом (редукция). Первому делению созревания предшествует 1 профаза мейоза. При сперматогенезе она протекает в 5 стадий: лептотена, зиготена,, пахитена, диплотена, диакинез.
Лептотене предшествует прелептотена, в этот период происходит удвоение ДНК (2n 4c). В лептотену хромосомы спирализуются и имеют вид тонких нитей. В зиготену происходит попарная конъюгация гомологичных хромосом и между ними происходит кроссинговер – обмен генными участками. Кроссинговер имеет важное генетическое значение, так как приводит к возникновению генетических различий между индивидуумами.
|
|
|
Пахитена – хромосомы продолжают спирализацию, они укорачиваются и утолщаются.
Диплотена – хромосомы входят в состав бивалентов, каждый состоит из 2 хромосом и каждая хромосома состоит из двух хроматид. Т.о. образуются тетрады, их количество 23, т.е. гаплоидное.
Диакинез – хромосомы утолщаются еще больше, их концы расходятся, они связаны перетяжкой и вступают в 1-ю метафазу мейоза. Происходит редукционное деление.
В результате из одного сперматоцита I возникает две равновеликие клетки - сперматоциты II порядка, они наследуют по 23 гомологичных хромосомы (1n2c).Затем наступает второе деление созревания. Оно протекает как обычный митоз, но при гаплоидном числе хромосом, т.е. удвоение ДНК не происходит. Такое деление называется эквационным («эквацио» - равенство), так как образуются две тождественные, т.е. полностью равноценные клетки, которые называются сперматидами. Всего образуется 4 сперматиды и каждая наследует по 23 хроматиды (1n1c)
|
В четвертом периоде - формирования - округлая сперматида приобретает форму зрелой мужской половой клетки: у нее вырастает жгутик, уплотняется ядро, происходит образование чехлика, уменьшается объем цитоплазмы, образуется оболочка, проиходит образование головки, шейки, хвоста. В результате всего процесса сперматогенеза из каждой исходной недифференцированной сперматогонии получается 4 зрелых половых клетки - сперматозоида, содержащих по гаплоидному набору хромосом.
Яйцеклетка и овогенез
Яйцеклетки, или овоциты (ovum), высокодифференцированные клетки, приспособленные к оплодотворению и дальнейшему развитию. Они отличаются от сперматозоидов однородной формой (преимущественно шаровидной, реже овальной) большими размерами, отсутствием центриолей. В связи с отсутствием центриолей яйцеклетки не способны к самостоятельному делению. Следует обратить внимание на то, что основным отличительным яйцеклеток различных представителей хордовых является количество желточных включений в цитоплазме. На основе количества и распределения желтка в цитоплазме яйцеклеток создана их классификация.
По количеству желтка различают следующие виды яйцеклеток.
|
|
|
1. Алецитальные (безжелтковые) - у видов, развитие которых протекает с метаморфозами и эмбриональный период очень короткий или у некоторых паразитарных червей.
2. Олиголецитальные (маложелтковые) - у видов развивающихся вне организма матери в относительно благоприятной водной среде, эмбриональный период относительно короткий (губки, иглокожие, круглоротые, ланцетник). А также у видов с внутриутробным развитием, зародыши которых питаются за счет матери (млекопитающие).
3. Мезолецитальные (среднее количество желтка) - развитие вне организма матери в водной среде (рыбы, земноводные) и у сумчатых млекопитающих.
4. Полилецитальные (многожелтковые) - развитие идет вне организма матери, причем на суше (птицы, пресмыкающиеся, яйцекладущие млекопитающие).
Таким образом, количество желтка в яйцеклетке зависит от условий, в которых развивается зародыш, а также в какой-то степени от длительности эмбрионального развития.
По распределению желтка в цитоплазме различают следующие виды яйцеклеток.
1. Изолецитальные - равномерное распределение желтка по всей цитоплазме. Характерно для олиголецитальных яйцеклеток. Различают первичноизолецитальные (ланцетник) и вторичноизолецитальные яйцеклетки (плацентарные млекопитающие).
2. Телолецитальные - желток распределяется по цитоплазме неравномерно, полярно. На одном полюсе (вегетативном) располагается желток, а на другом полюсе (анимальном) - ядро и органоиды. Характерно для мезо- и полилецитальных яйцеклеток (земноводные, птицы, яйцекладущие и сумчатые млекопитающие). Среди телолецитальных яйцеклеток различают 2 подгруппы:
а) умеренно телолецитальные - полярность выражено умеренно, нерезко (мезолецитальная яйцеклетка лягушки);
б) резко телолецитальные - полярность ярко выражена (полилецитальная яйцеклетка рыбы, птицы).
3. Центролецитальные - желток в виде узкого пояска сосредоточен вокруг ядра (насекомые).
Таким образом, у ланцетника яйцеклетка олиголецитальная и первично- изолецитальная, у лягушки - мезолецитальная и умеренно телолецитальная, у птиц - полилецитальная и резко телолецитальная, у млекопитающих - олиголецитальная и вторично изолецитальная.
|
|
|
Верхняя часть яйцеклетки, где мало желтка называется анимальным полюсом, нижняя, где желтка больше – вегетативным полюсом.
Яйцеклетки имеют следующие оболочки:
первичную - собственная оболочка (оолемма);
вторичную - продукт деятельности самой яйцеклетки и соседних вспомогательных клеток (например, фолликулярных);
третичную - имеется у видов развивающихся вне организма матери на суше, и является продуктом деятельности слизистой оболочки яйцевыводящих путей.
Процесс развития женских половых клеток называется овогенезом. В отличие от сперматогенеза он имеет некоторые особенности.
В овогенезе различают 3 периода: размножение, рост и созревание. Развитие женских половых клеток происходит в яичниках. Период размножения наступает у оогонии еще у зародыша и прекращается к моменту рождения девочки.
Недифференцированные женские половые клетки - овогонии -размножаются так же, как и сперматогонии, путем обычного митоза. После деления они становятся овоцитами I порядка и переходят в период роста.
Рост овоцитов длится очень долго - недели, месяцы и даже годы. В периоде роста различают два этапа: малый, или медленный рост, когда ассимилируются новые вещества и ими обогащается преимущественно цитоплазма, и большой, или быстрый рост, когда в клетке накапливается питательное вещество желток, который будет необходим в дальнейшем для первых дроблений зиготы. В фазе малого роста происходит образование большого количества разных типов РНК. Глубокие изменения в периоде роста претерпевает и ядро, оно сильно набухает, содержимое его как бы расплывается. Размеры клеток колоссально возрастают (например, икринки окуня увеличиваются почти в миллион раз). Впериод роста яйцеклетка проходит 1 профазу мейоза. Она включает все стадии, кроме диакинеза. С овоцитом 1 порядка происходят аналогичные изменения, которые сопровождают сперматоцит на аналогичной стадии (удвоение ДНК, спирализация и конъюгация хромосом, кроссинговер)
Затем овоцит I порядка вступает в период созревания, или мейоз. Здесь тоже совершаются редукционное и эквационное деления. Процессы деления в ядре протекают так же, как при мейозе сперматоцитов, но судьба цитоплазмы совершенно иная. При редукционном делении одно ядро увлекает с собой большую часть цитоплазмы, а на долю другого остается лишь незначительная ее часть. Поэтому образуется только одна полноценная клетка - овоцит II порядка, и вторая крошечная - направительное, или редукционное, тельце, которое может делиться на два редукционных тельца. Объем генетической информации клетки получают одинаковый (1n2c).
|
|
|
При втором, эквационной делении несимметричное распределение цитоплазмы повторяется и опять образуется одна крупная клетка - овотида и третье полярное тельце. Овотида по составу ядра и функционально является вполне зрелой половой клеткой. Все клетки получают гаплоидный набор сестринских хроматид (1n1c). В результате при оогенезе получается 4 клетки, из которых только одна станет в дальнейшем яйцеклеткой, а остальные 3 (полярные тельца) редуцируются. Биологическая значимость этого этапа оогенеза - сохранить все накопленные вещества цитоплазмы около одного гаплоидного ядра для обеспечения нормального питания и развития оплодотворенной яйцеклетки.
Период формирования, в отличие от сперматогенеза, в овогенезе отсутствует. Таким образом, в овогенезе из одной овогонии возникает только одна зрелая яйцеклетка. Полярные тельца остаются недоразвитыми и вскоре погибают и фагоцитируются другими клетками. Зрелые женские гаметы называют яйцеклетками или яйцами, а отложенные в воду - икрой.
Отличительные особенности сперматогенеза и овогенеза у человека представлены в таблице 3.
Табл.3
Таблица 3
| № п/п | Сперматогенез | Овогенез |
| 1. | Фаза размножения происходит только с момента полового созревания и продолжается в течение всей жизни | Фаза размножения происходит только в эмбриональном периоде и непродолжительное время после рождения |
| 2. | Фаза роста сразу следует за размножением, короткая | Фаза роста очень длительная, делится на малый рост и большой рост |
| 3. | Фаза созревания характеризуется равномерным делением сперматоцитов | Фаза созревания характеризуется неравномерным делением овоцитов: образуется одна яйцеклетка и три редукционных тельца |
| 4. | Есть фаза формирования | Фаза формирования отсутствует |
| 5. | «Экономичность» сперматогенеза: из одной сперматогонии образуется 4 сперматозоида | «Расточительность» овогенеза: из одной овогонии образуется одна крупная яйцеклетка и 3 мелких редукционных тельца |
| 6. | Продолжается в течение всей жизни мужчины | Прекращается после менопаузы |
Отличия половых клеток от соматических
1. Набор хромосом у половых клеток гаплоидный, у соматических клеток - диплоидный.
2. Для половых клеток характерно сложное, стадийное развитие; при этом имеет место особый способ деления - мейоз.
3. Половые клетки имеют специальные приспособления: - сперматозоид имеет акросому (для проникновения через оболочки яйцеклетки) и мощный двигательный аппарат - хвостик; яйцеклетка имеет желток (запас питательных веществ и строительных материалов) и оболочки (I, II, а у некоторых видов и III).
4. У половых клеток особое ядерно-цитоплазматическое отношение: у мужских половых клеток очень высокое (преобладает ядро над цитоплазмой), в женских половых клетках очень низкое (преобладает цитоплазма над ядром).
5. Обмен веществ в зрелых половых клетках до оплодотворения находится на очень низком уровне (почти до анабиоза).
6. Биологическое назначение: если из соматической клетки может образоваться лищь такая же дочерняя клетка, то из половых клеток формируется целый новый организм.
Оплодотворение
Оплодотворение - процесс слияния мужской и женской половых клеток, в результате чего восстанавливается диплоидный набор хромосом, характерный для того или иного вида животных, и возникает качественно новая клетка - зигота.
Встреча гамет происходит либо внутри половых путей самки {внутреннее оплодотворение), либо во внешней среде, например, в воде (наружное оплодотворение).
Сперматозоид приближается к яйцеклетке головкой вперед. В случае если оболочка яйцеклетки мягкая, навстречу ему приподнимается протоплазматический вырост яйца - воспринимающий бугорок, который и втягивает спермий в глубь яйца. После этого почти мгновенно над воспринимающим бугорком появляется тонкая желточная оболочка оплодотворения, наглухо закрывающая сюда доступ остальным спермиям. При плотных оболочках спермин проникают в яйцеклетки через одно из микропилярных отверстий. В процессе оплодотворения различают три фазы. Последовательность стадий оплодотворения представлена на рис. 6.
При оплодотворении различают следующие фазы.
Первая фаза – дистатные взаимодействия гамет или сближение. Как при наружном (у рыб, амфибий), так и при внутреннем (у рептилий, птиц и млекопитающих) оплодотворении сперматозоиды в результате хемотаксиса в условиях слабо щелочной среды очень быстро перемещаются по направлению к яйцеклеткам. Смещение рН в кислую сторону, наоборот, парализует спермин. Сперматозоиды млекопитающих обладают также реотаксисом, т.е. способностью двигаться против тока жидкости. Яйцеклетка и сперматозоид выделяют особые химические вещества - гамоны, которые обеспечивают им встречу: гиногамоны –(гормоны яйцеклетки) и андрогомоны (гормоны сперматозоида). Гиногамоны бывают 2-х видов: гиногамоны-1(небелковой природы, выделяются яйцевыми оболочками) - стимулируют движение сперматозоида к яйцеклетке, а гиногамоны-2 (белково-углеводной природы: фертилизины, агглютинины,) – вызывают агглютинацию (склеивание) сперматозоидов и тем самым защищают яйцеклетку от проникновения в нее избыточного количества сперматозоидов.
Сперматозоиды также выделяют: андрогомон-1 (небелковой природы) – антагонист гиногамона-1, подавляет подвижность сперматозоидов и спермолизины акросомы – растворяют оболочки яйцеклетки при контактных взаимодействиях.
Вторая фаза – контатные взаимодействия гамет или проникновение сперматозоида через оболочки яйцеклетки. Контактное взаимодействие гамет наступает, когда сперматозоид сближается с яйцеклеткой. У млекопитающих при оплодотворении в яйцеклетку проникает лишь один сперматозоид. Такое явление называется моноспермией. У беспозвоночных животных, рыб, амфибий, рептилий и птиц возможна полиспермия, когда в яйцеклетку проникает несколько сперматозоидов, но в слиянии ядер (оплодотворении) все равно принимает участие только один. Различают акросомную и кортикальную реакции. Акросомная реакция - это реакция сперматозоида на контакт не только с яйцеклеткой, но и с любой твердой поверхностью. В месте контката со сперматозоидом яйцеклетка образуетс воспринимающий бугорок. В течение короткого времени (10-20 сек.) происходят измененимя акросомного аппарата. Наружная мембрана акросомы разрывается и высвобождаются ферменты (спермолизины), они быстро растворяют облочки яйцеклетки в месте контакта. Внутренняя оболочка акросомы образует вырост – трубочку, которая растет и срастается с плазмалеммой яйцеклетки. Образуется цитоплазматический мостик, через который ядро, клеточный центр сперматозоида переходят в яйцеклетку. Мембрана сперматозоида встраивается в мембрану яйцеклетки.
Проникновение сперматозоида значительно усиливает процессы внутриклеточного обмена, что связано с повышением дыхания и активизацией ферментативных систем яйцеклетки. Развивается кортикальная реакция. После соприкосновения со сперматозоидом изменяется величина мембранного потенциала, происходит внутриклеточное высвобождение, повышение, а затем понижение концентрации ионов кальция в цитоплазме. После этого начинается экзоцитоз кортикальных гранул (располагаются в периферическом слое цитоплазмы яйцеклетки), который распространяется от места контакта со сперматозоидом во все стороны. Вещества, которые высвобождаются из кортикальных гранул приводят к расслоению между плазмалеммой и желточной оболочкой или оболочкой оплодотворения. Последняя отслаивается и быстро затвердевает. Между плазмалеммой и желточной оболочкой образуется перивителлиновое пространство, оно заполняется содержимым кортикальных гранул. Кортикальная реакция препятствует проникновению в женскую половую клетку избытка сперматозоидов, они приклеиваются к желточной оболочке

Рис. 6. Схематическое изображение процесса оплодотворения (В.Г.Влисеев, 1983): 1 - цитоплазма яйцеклетки; 2 - ядро яйцеклетки; 3 - блестящая оболочка; 4 - фолликулярный эпителий; 5 - головка сперматозоида; 6 - шейка сперматозоида; 7 - хвост сперматозоида; 8 - воспринимающий бугорок; 9 - оболочка оплодотворения; 10 - женский пронуклеус; 11 - мужской пронуклеус; 12 - веретено между центриолями; 13 - синкарион
Третья фаза – сингамия - образование мужского и женского пронуклеусов с последующим слиянием их (синкарион). При этом у многих видов животных ядра мужской и женской клеток во время сближения переходят в состояние метафазы. Затем хромосомы обоих ядер образуют единую материнскую «звезду», но уже с удвоенным (диплоидным) числом хромосом. В других случаях ядра вначале сливаются и затем переходят в состояние кариокинеза. Одновременно внесенные сперматозоидом центриоли расходятся к полюсам клетки, и этот одноклеточный зародыш - зигота вступает во второй период эмбрионального развития - период дробления.
К занятию №3
Дробление
Дробление - это последовательные митотические деления зиготы, в результате чего зародыш становится многоклеточным существенно не изменяя своего объема. При дроблении интерфазы между делениями короткие, поэтому бластомеры не успевают расти, и с каждым делением дробления становятся все меньше и меньше, при этом количество бластомеров увеличивается, а объем каждого отдельного бластомера уменьшается. Дочерние клетки называются бластомерами, они не расходятся. Плоскости по которым происходят деления называются бороздами дробления. Борозды дробления бывают меридианальные, широтные (латитудинальные), экваториальные и тангенциальные.
Меридианальные – проходят от полюса к полюсу.
Экваториальные – проходят в центре бластулы.
Широтные – выше и ниже экватора.
Тангенциальные – идут параллельно касательной.
Тип дробления зависит от типа яйцеклетки, т.е. от количества и распределения желтка, а также от взаимного расположения дробящихся клеток.
Выделяют следующие типы дробления зиготы.
Полное дробление - голобластическое (holos - весь, blastos - зачаток) - в дроблении участвуют все участки зародыша. Это деление может быть:
равномерным (синхронным) - когда все бластомеры дробятся с одинаковой скоростью и поэтому количество их увеличивается в геометрической прогрессии, т.е. происходит кратное увеличение бластомеров (1, 2, 4, 8 и т.д.). Характерно для яйцеклеток с малым количеством желтка, при этом образуются бластомеры примерно одинакового размера (ланцетник);
неравномерным (асинхронным) - когда количество бластомеров увеличивается в неправильной прогрессии (1, 2, 3, 5 и т.д.). Характерно для яйцеклеток со средним содержанием желтка (круглоротые, хрящевые рыбы, земноводные). При этом образуются бластомеры неодинакового размера. Сначала в результате первых двух дроблений образуются бластомеры примерно одинакового размера, а затем на анимальном полюсе деление происходит быстрее, чем на вегетативном. В результате на анимальном полюсе образуется большее количество бластомеров, и они меньшего размера, чем на вегетативном полюсе. В дальнейшем эти бластомеры дифференцируются по-разному - из одних образуется тело зародыша, а другие выполняют трофическую функцию.
Неполное дробление (частичное) - меробластическое - дробление идет только на анимальном полюсе, вегетативный полюс перегружен желтком и в дроблении не участвует. Это дробление может быть:
поверхностное - дробится поверхностная часть зиготы, а центральная часть, богатая желтком не делится (членистоногие);
дискоидальное - дробится небольшой участок зиготы, где мало желтка, а остальная часть, богатая желтком, не делится (костистые рыбы, пресмыкающиеся, птицы).
В зависимости от расположения делящихся клеток различают три типа дробления:
радиальное - когда верхний ряд бластомеров располагается точно над нижним рядом (кишечнополостные, иглокожие, низшие хордовые);
спиральное - когда верхний ряд бластомеров располагается между клетками нижнего ряда (большинство червей, моллюски);
двусимметричное (билатеральное)- когда делящиеся клетки располагаются симметрично по бокам от исходного бластомера (круглые черви, асцидии);
анархическое - отсутствие закономерности в расположении бластомеров у организмов одного вида.
В процессе деления зиготы часто сочетаются различные типы дробления. В процессе дробления развивающийся зародыш проходит последовательно три стадии развития - бластула, гаструла, нейрула.
|
|
|
