 |


Кислородная емкость крови. Механизм транспорта кислорода кровью. Анализ кривой диссоциации оксигемоглобина.
|
|
|
|
Транспорт О2 осуществляется в физически растворенном и химически связанном виде. Физические процессы, т. е. растворение газа, не могут обеспечить запросы организма в О2. Подсчитано, что физически растворенный О2 может поддерживать нормальное потребление О2 в организме (250 мл*мин-1), если минутный объем кровообращения составит примерно 83 л*мин-1 в покое. Наиболее оптимальным является механизм транспорта О2 в химически связанном виде.
Согласно закону Фика, газообмен О2 между альвеолярным воздухом и кровью происходит благодаря наличию концентрационного градиента О2 между этими средами. В альвеолах легких парциальное давление О2 составляет 13,3 кПа, или 100 мм рт.ст., а в притекающей к легким венозной крови парциальное напряжение О2 составляет примерно 5,3 кПа, или 40 мм рт.ст. Давление газов в воде или в тканях организма обозначают термином «напряжение газов» и обозначают символами Ро2, Рсo2. Градиент О2 на альвеолярно-капиллярной мембране, равный в среднем 60 мм рт.ст., является одним из важнейших, но не единственным, согласно закону Фика, факторов начальной стадии диффузии этого газа из альвеол в кровь.
Транспорт О2 начинается в капиллярах легких после его химического связывания с гемоглобином.
Гемоглобин (Нb) способен избирательно связывать О2 и образовывать оксигемоглобин (НbО2) в зоне высокой концентрации О2 в легких и освобождать молекулярный О2 в области пониженного содержания О2 в тканях. При этом свойства гемоглобина не изменяются и он может выполнять свою функцию на протяжении длительного времени.
Гемоглобин переносит О2 от легких к тканям. Эта функция зависит от двух свойств гемоглобина: 1) способности изменяться от восстановленной формы, которая называется дезоксигемоглобином, до окисленной (Нb + О2 à НbО2) с высокой скоростью (полупериод 0,01 с и менее) при нормальном Рог в альвеолярном воздухе; 2) способности отдавать О2 в тканях (НbО2 à Нb + О2) в зависимости от метаболических потребностей клеток организма.
|
|
|
Зависимость степени оксигенации гемоглобина от парциального давления Ог в альвеолярном воздухе графически представляется в виде кривой диссоциации оксигемоглобина, или сатурационной кривой (рис. 8.7). Плато кривой диссоциации характерно для насыщенной О2 (сатурированной) артериальной крови, а крутая нисходящая часть кривой — венозной, или десатурированной, крови в тканях.
На сродство кислорода к гемоглобину влияют различные метаболические факторы, что выражается в виде смещения кривой диссоциации влево или вправо. Сродство гемоглобина к кислороду регулируется важнейшими факторами метаболизма тканей: Ро2 pH, температурой и внутриклеточной концентрацией 2,3-дифосфоглицерата. Величина рН и содержание СО2 в любой части организма закономерно изменяют сродство гемоглобина к О2: уменьшение рН крови вызывает сдвиг кривой диссоциации соответственно вправо (уменьшается сродство гемоглобина к О2), а увеличение рН крови — сдвиг кривой диссоциации влево (повышается сродство гемоглобина к О2) (см. рис. 8.7, А). Например, рН в эритроцитах на 0,2 единицы ниже, чем в плазме крови. В тканях вследствие повышенного содержания СО2 рН также меньше, чем в плазме крови. Влияние рН на кривую диссоциации оксигемоглобина называется «эффектом Бора».
Рост температуры уменьшает сродство гемоглобина к О2. В работающих мышцах увеличение температуры способствует освобождению О2. Уменьшение температуры тканей или содержания 2,3-дифосфоглицерата вызывает сдвиг влево кривой диссоциации оксигемоглобина (см. рис. 8.7, Б).
Метаболические факторы являются основными регуляторами связывания О2 с гемоглобином в капиллярах легких, когда уровень O2, рН и СО2 в крови повышает сродство гемоглобина к О2 по ходу легочных капилляров. В условиях тканей организма эти же факторы метаболизма понижают сродство гемоглобина к О2 и способствуют переходу оксигемоглобина в его восстановленную форму — дезоксигемоглобин. В результате О2 по концентрационному градиенту поступает из крови тканевых капилляров в ткани организма.
|
|
|
Оксид углерода (II) — СО, способен соединяться с атомом железа гемоглобина, изменяя его свойства и реакцию с О2. Очень высокое сродство СО к Нb (в 200 раз выше, чем у О2) блокируют один или более атомов железа в молекуле гема, изменяя сродство Нb к О2.
Под кислородной емкостью крови понимают количество Ог, которое связывается кровью до полного насыщения гемоглобина. При содержании гемоглобина в крови 8,7 ммоль*л-1 кислородная емкость крови составляет 0,19 мл О2 в 1 мл крови (температура 0oC и барометрическое давление 760 мм рт.ст., или 101,3 кПа). Величину кислородной емкости крови определяет количество гемоглобина, 1 г которого связывает 1,36—1,34 мл О2. Кровь человека содержит около 700—800 г гемоглобина и может связать таким образом почти 1 л О2. Физически растворенного в 1 мл плазмы крови О2 очень мало (около 0,003 мл), что не может обеспечить кислородный запрос тканей. Растворимость О2 в плазме крови равна 0,225 мл*л-1*кПа-1
Обмен О2 между кровью капилляров и клетками тканей также осуществляется путем диффузии. Концентрационный градиент О2 между артериальной кровью (100 мм рт.ст., или 13,3 кПа) и тканями (около 40 мм рт.ст., или 5,3 кПа) равен в среднем 60 мм рт.ст. (8,0 кПа). Изменение градиента может быть обусловлено как содержанием О2 в артериальной крови, так и коэффициентом утилизации О2, который составляет в среднем для организма 30— 40%. Коэффициентом утилизации кислорода называется количество О2, отданного при прохождении крови через тканевые капилляры, отнесенное к кислородной емкости крови.
Билет 11
1. Мембрана представляет собой двойной липидный слой, в который погружены интегральные белки, функционирующие как ионные насосы и каналы.Насосы за счёт энергии АТФ перекачивают ионы K, Na, Ca против градиента концентрации. Периферические белки формируют цитоскелет клетки, который придаёт прочность и в тоже время эластичность клетке. Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды и холестерол. Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим — более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку. Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются.Клеточная мембрана является разделительным барьером между цитоплазмой и внеклеточной средой. Транспорт веществ сквозь клеточную мембрану в клетку или из клетки, осуществляемый с помощью различных механизмов — простой диффузии, облегченной диффузии и активного транспорта. Важнейшее свойство биологической мембраны состоит в ее способности пропускать в клетку и из нее различные вещества. Это имеет большое значение для саморегуляции и поддержания постоянного состава клетки. Такая функция клеточной мембраны выполняется благодаря избирательной проницаемости, то есть способностью пропускать одни вещества и не пропускать другие.
|
|
|
В клетке существует 4 основных вида транспорта: 1) Диффузия, 2) Осмос, 3) Активный транспорт, 4) эндо и экзоцитоз. 1) Диффузия — это перемещение веществ по диффузному градиенту, т.е. из области высокой концентрации, в область с низкой концентрацией. Медленно диффундируют ионы, глюкоза, аминокислоты, липиды и т.д. Быстро диффундируют жирорастворимые молекулы. Облегченная диффузия является модификацией диффузии. Наблюдается в том случае, когда определенному веществу помогает пройти через мембрану какая-либо специфическая молекула, т.е. у этой молекулы есть свой канал, через который она легко проходит (поступление глюкозы в эритроциты). 2) Осмос — это дифундированние воды через полупроницаемые мембраны. 3) Активный — это перенос молекул или ионов через мембрану, против градиента концентрации и электрохимического градиента.белки-переносчики (их иногда называют белки-насосы) переносят через мембрану вещества с затратами энергии, которая обычно поставляется при гидролизе АТФ. В клетке между двумя сторонами плазматической мембраны поддерживается разность потенциалов — мембранный потенциал. Внешняя среда положительный заряд, а внутренняя отрицательный. Поэтому в клетку будут стремится катионы Na, K, а анионы хлора будут отталкиваться. Примером активного транспорта имеющегося в большинстве клеток является натриево-калиевый насос. 4) Эндо и экзоцитоз. Плазматическая мембрана принимает учатие в выведении веществ из клетки, это происходит в процессе экзоцитоза. Так выводятся гормоны, полисахариды, белки, жировые капли и др. продукты клетки. Они заключаются в пузырьки, ограниченные мембраной, и подходят к плазмолеме. Обе мембраны сливаются и содержимое пузырька выводится наружу. Фагоцтоз - захват и поглощение клеткой крупных частиц. Пиноцитоз — процесс захвата и поглощения капелек жидкости.
|
|
|
Калий/натриевый насос. Первоначально этот переносчик присоединяет с внутренней стороны мембраны три иона 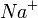 . Эти ионы изменяют конформацию активного центра АТФазы. После такой активации АТФаза способна гидролизовать одну молекулу АТФ, причем фосфат-ион фиксируется на поверхности переносчика с внутренней стороны мембраны.Выделившаяся энергия расходуется на изменение конформации АТФазы, после чего три иона и ион
. Эти ионы изменяют конформацию активного центра АТФазы. После такой активации АТФаза способна гидролизовать одну молекулу АТФ, причем фосфат-ион фиксируется на поверхности переносчика с внутренней стороны мембраны.Выделившаяся энергия расходуется на изменение конформации АТФазы, после чего три иона и ион  (фосфат) оказываются на внешней стороне мембраны. Здесь ионы отщепляются, а замещается на два иона
(фосфат) оказываются на внешней стороне мембраны. Здесь ионы отщепляются, а замещается на два иона 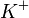 . Затем конформация переносчика изменяется на первоначальную, и ионы оказываются на внутренней стороне мембраны. Здесь ионы отщепляются, и переносчик вновь готов к работе.
. Затем конформация переносчика изменяется на первоначальную, и ионы оказываются на внутренней стороне мембраны. Здесь ионы отщепляются, и переносчик вновь готов к работе.
Более кратко действия АТФазы можно описать так:
4. 1) Она изнутри клетки «забирает» три иона ,затем расщепляет молекулу АТФ и присоединяет к себе фосфат
5. 2) «Выбрасывает» ионы и присоединяет два иона из внешней среды.
6. 3) Отсоединяет фосфат, два иона выбрасывает внутрь клетки
В итоге во внеклеточной среде создается высокая концентрация ионов , а внутри клетки — высокая концентрация . Работа , — АТФаза создает не только разность концентраций, но и разность зарядов (она работает как электрогенный насос). На внешней стороне мембраны создается положительный заряд, на внутренней — отрицательный.
|
|
|
|
|
|
