 |


Восход нуклеиновой кислоты
|
|
|
|
Если у биохимиков и оставались еще какие-то былые сомнения в том, что нести характеристики всего живого может нечто иное, нежели белок, то эксперименты, проведенные в начале 1950-х годов на материале вирусов, окончательно их развеяли.
После Второй мировой войны производство электронных микроскопов дошло до такого уровня, когда этот инструмент стал, во-первых, вполне надежным, а во-вторых, доступным по цене для средней научной лаборатории. Электронные микроскопы, чьи увеличительные возможности в значительной степени превышали возможности обычных оптических микроскопов, впервые позволили воочию лицезреть вирусы, что резко увеличило интерес к их изучению.
Оказалось, что вирусы состоят обычно из полой белковой оболочки, внутри которой скрывается молекула нуклеиновой кислоты. Последняя представляет собой единую длинную цепь, а белковая оболочка — последовательность одинаковых небольших секций. На этом этапе стало вдруг понятно, что белок в общем-то не столь уж обязательно является более сложным соединением, чем нуклеиновая кислота. Вирусы явили миру пример структуры, в которой молекулы нуклеиновой кислоты
были явно крупнее любой имеющейся в ней белковой молекулы. Разумеется, сам по себе размер еще не означает сложности, о чем я уже упоминал и к чему еще вернусь.
В 1952 году двое биохимиков, Альфред Херши и Марта Чейз, провели один крайне важный эксперимент на бактериофагах — вирусах, заражающих клетки бактерий. Эти вирусы проникают в клетку и начинают размножаться в ней до тех пор, пока клетка не умирает. Клеточная мембрана лопается, и вирусы разлетаются наружу.
Херши и Чейз сначала вырастили бактерии в среде, содержащей радиоактивные атомы серы и фосфора. С химической точки зрения такие атомы ведут себя, как обычные атомы серы и фосфора, так что бактерии обычным образом поглощали их и встраивали в состав собственной ткани. Однако радиоактивные атомы отличаются от обычных тем, что испускают энергонесущие частицы, которые с помощью специальной аппаратуры можно отследить. Причем излучение радиоактивного фосфора отличается от излучения радиоактивной серы. Иными словами, выросшие в такой радиоактивной среде бактерии становятся «мечеными».
|
|
|
На втором этапе эти бактерии были заражены бактериофагами. И вирусы принялись размножаться в клетках, используя для собственного строительства материалы клетки — то есть новые вирусы тоже оказывались мечеными. Однако в метках вирусов наблюдалась особая закономерность. Дело в том, что в белковых молекулах практически неизбежно имеется атом серы, а фосфор есть далеко не всегда, да и то в крайне небольших количествах. А вот в состав нуклеиновых кислот, как раз наоборот, фосфор входит всегда, а сера — никогда. То есть если бактериофаг несет и «фосфорную», и «серную» метки, то имеющийся в нем фосфор находится в ядре, состоящем из нуклеиновой кислоты, а сера — в белковой оболочке.
И вот настал последний, решающий этап. Мечеными бактериофагами были заражены нормальные, нерадиоактивные бактерии. Теперь наличие радиоактивных атомов могло свидетельствовать только о присутствии вируса. Так вот, в бактерии внедрялся только один радиоактивный элемент из двух. И это был фосфор. А радиоактивная сера оставалась снаружи, и ее спокойно можно было смыть, или даже просто стряхнуть.
Неизбежно напрашивался вывод, что в бактерию проникала лишь сердцевина вируса — нуклеиновая кислота, а белковая оболочка оставалась снаружи, сброшенной. А оказавшись внутри бактерии, нуклеиновая кислота вируса быстро начинала строить не только подобные ей молекулы нуклеиновой кислоты, отличные от нуклеиновых кислот, свойственных самой бактерии, но также и новые белковые оболочки!
|
|
|
Теперь уже никуда нельзя было деться от того факта, что носителем генетического кода, по крайней мере в данном конкретном случае, является не белок, а нуклеиновая кислота; что именно нуклеиновая кислота, без помощи белка, способна управлять формированием специфических белковых молекул. По крайней мере, молекулы белковой оболочки новых вирусов были в точности такими же, как и те, что остались сброшенными перед внедрением в бактерию, отличаясь от всех белков, имевшихся в самой бактерии.
Через несколько лет по белковой теории наследственности был нанесен следующий удар. В 1955 году Хайнц Френкель-Конрат разработал тонкие технологии выделения нуклеиновой кислоты из белковой оболочки вируса табачной мозаики без вреда как для белка, так и для самой нуклеиновой кислоты. По отдельности каждая из обеих составляющих не имела инфицирующей способности — то есть и белок, и кислоту можно было распылять на листья табака без риска вызвать само заболевание, которое сразу стало бы заметно по своим симптомам — характерному крапчатому обесцвечиванию листьев. Однако если снова смешать разделенные белок и нуклеиновую кислоту, то часть молекул последней каким-то образом вновь попадала обратно в белковую оболочку и получившаяся смесь опять обретала способность к заражению растений. На следующий год Френкель-Конрат сумел уточнить, что это только белок полностью теряет инфицирующую способность после разделения вируса, а нуклеиновая кислота все же сохраняет ее, хотя и в крайне малых объемах.
Вывод ясен. Белковая оболочка служит, в первую очередь, как «скелет», предназначение которого — защищать главную, нуклеиновокислую, часть вируса. Кроме того, в белковой оболочке содержится фермент, проделывающий в клетке бактерии входное отверстие (сам фермент в итоге был выделен в 1962 году), через которое проникает нуклеиновая кислота, отбросив отработавший свое белок.
В отсутствие белковой оболочки нет и фермента, который бы открыл нуклеиновой кислоте путь в клетку, поэтому чистая нуклеиновая кислота теряет способность заражать подавляющее большинство клеток. Сама по себе она может проникнуть в клетку только через какую-нибудь случайную трещину, и тогда происходит полноценное заражение даже при отсутствии белка.
|
|
|
Можно провести аналогию между нуклеиновой кислотой в белке и человеком в автомобиле. Человек в автомобиле без проблем доберется от Нью-Йорка до Чикаго. Если же человека и автомобиль разделить, задача эта окажется для каждого из них по отдельности практически невыполнимой. Автомобиль сам по себе вообще никуда не сдвинется, человек же, конечно, может дойти от Нью-Йорка до Чикаго пешком, но для того у него должны быть ну очень серьезные мотивы. В целом же понятно, кто из пары «автомобиль и человек» является главным, и так же ясно, что нуклеиновая кислота — главный член пары «белок и нуклеиновая кислота».
Все эксперименты, проводимые начиная с 1944 года, указывали на то же самое. Именно нуклеиновая кислота является носителем генетического кода всех видов живых существ, и клеток, и вирусов. Белок никогда не несет этой информации. Так что с конца 1940-х годов химики переключили свой интерес на молекулы нуклеиновых кислот.
То же самое предстоит сделать и нам, поскольку сейчас мы начнем разбираться в строении нуклеиновой кислоты (как до этого разбирались в строении белков), для того чтобы проникнуть в природу генетического кода.
Глава 7
«ЗОЛУШКА»
ФОСФОР
На момент обретения в 1944 году всемирной славы нуклеиновая кислота была уже известна химикам на протяжении трех четвертей века. Однако лишь немногие удосужились за все это время хоть немного поизучать ее. Это была «Золушка», жившая тихой и неприметной бедной родственницей среди более интересных химических соединений, пока вдруг не обнаружилось, что именно ей подходит тот самый хрустальный башмачок.
И все же тем немногим первопроходцам, которые работали над нуклеиновой кислотой и ранее, удалось установить важнейшие особенности ее строения. Так, уже вскоре после ее открытия стало известно, что в состав нуклеиновой кислоты входит фосфор.
|
|
|
Это многих удивило. Известно было, что некоторые белки тоже содержат фосфор, но в очень небольших количествах. Казеин — основной белок молока, содержит 1 процент фосфора. Лецитин, жирное вещество, находящееся в яичном желтке, состоит из фосфора на 3 процента. А в нуклеиновой кислоте этого элемента содержится более, чем в любом другом веществе организма, — 9 процентов!
Пора, наверное, поподробнее остановиться на том, что представляет из себя фосфор. Как уже упоминалось в главе 3, в химических формулах он обозначается буквой Р. По химическим свойствам фосфор в какой-то мере похож на азот. Как и азот, он имеет три валентности, а кроме того, в некоторых случаях может присоединять к себе еще и четвертый атом (как правило, это оказывается кислород) — с помощью особого вида связи, на схемах обозначаемого стрелочкой[21]. Пример такой связи вы можете видеть на рис. 35, где изображена структурная формула фосфорной кислоты — широко используемого в химической промышленности вещества. Эмпирическая формула его, как видите, Н2РO4. Обратите внимание, что атом кислорода, обычно образующий две связи «стандартного» типа, «стрелочную» связь образует только одну.
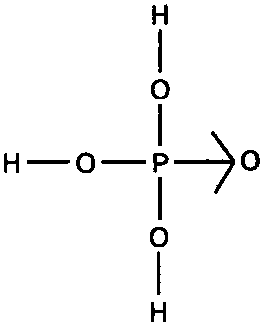
Рис. 35. Фосфорная кислота
Связи между атомами фосфора и кислорода в фосфорной кислоте очень прочны, а вот атомы водорода достаточно легко можно из нее «выковырять». Если это сделать, то у оставшейся части фосфорной кислоты появится свободная валентность для присоединения других атомов или групп атомов. Если удалить два атома водорода, появится две свободные валентности, если три — то три. Фосфорную кислоту, лишенную одного или нескольких атомов водорода, называют фосфатной группой. В зависимости от количества устраненных атомов водорода различают первичные, вторичные и третичные фосфаты — они показаны на рис. 36.

Рис. 36. Фосфатные группы
В живой ткани атомы фосфора представлены только в составе первичных или вторичных фосфатных групп. Поэтому в целях упрощения изложения будет лучше просто обозначать эти две группы стандартным образом, не вдаваясь в их внутреннее атомное строение. Удобно будет ввести особый знак — обведенную окружностью букву Р для обозначения не атома фосфора, а фосфатной группы. Различать между собой первичные и вторичные фосфаты мы будем, указывая одну или две валентности у единого символа, как на рис. 37.

Рис. 37. Обозначения фосфатных групп
В качестве примера того, в каком виде фосфатная группа может встречаться в сочетании с уже рассмотренными нами соединениями, давайте рассмотрим аминокислоту серин. В некоторых случаях фосфатная группа крепится к серину вместо кислорода в радикале. Тогда мы получаем фосфосерин (см. рис. 38).
|
|
|

Рис. 38. Серин и фосфосерин
Фосфосерин иногда встречается в белках на месте серина, — получается фосфорсодержащий белок. Примером такого белка является упомянутый мной в начале главы казеин.
Так что запишем в список составляющих нуклеиновой кислоты фосфатные группы и отметим, что именно их присутствие придает всему соединению кислотные свойства. Но разумеется, ими нуклеиновая кислота не ограничивается.
ДВЕ РАЗНОВИДНОСТИ
Практически с самого начала изучения нуклеиновой кислоты отмечалось, что в нее входят сахарные группы, но конкретная природа этих групп на протяжении десятилетий оставалась загадкой.
Самый распространенный в природе, простой сахар — это глюкоза, тот самый материал, из которого строятся крахмал и целлюлоза. Молекула глюкозы представляет собой цепочку из шести атомов углерода, к пяти из которых прикреплены гидроксильные группы, а шестой входит в состав карбонильной группы. Именно наличие карбонильной группы и нескольких гидроксильных и является характеризующей особенностью строения сахаров.
Также в числе распространенных сахаров можно назвать фруктозу и галактозу. В их основе тоже лежит цепочка из шести атомов углерода, один из которых входит в карбонильную группу, а остальные связаны с гидроксильными группами. Различаются они относительной ориентацией гидроксильных групп в пространстве — этого вполне достаточно для того, чтобы соединения имели различные химические свойства.
Два простых сахара могут, как и аминокислоты, объединяться между собой с выделением воды. Соединение глюкозы и фруктозы называется сахарозой — это и есть тот самый столовый сахар, который мы кладем в чай и кофе. И кленовый сахар, и свекловичный сахар, и тростниковый сахар — все это сахароза. А соединение глюкозы с галактозой называется лактозой, этот сахар почти безвкусен и встречается только в молоке. Ну и, как мы знаем, объединяясь между собой, молекулы глюкозы формируют крахмал или целлюлозу.
Существует множество и других сахаров и их сочетаний, так же как и видоизмененных сахаров, где к сахарным молекулам присоединены азото-, серо- или фосфорсодержащие группы. Некоторые из подобных соединений в природе не встречаются, но тем не менее создаются в лабораторных условиях.
Все эти вещества — простые, сложные, или комбинированные, натуральные или синтетические — имеют общее название углеводы. Как уже отмечалось в главе 2, это одна из трех основных групп, на которые делят органические вещества, обнаруживаемые в живых тканях.
Но какой именно углевод содержится в нуклеиновой кислоте? Ответа на этот вопрос не было до 1910 года, когда американский биохимик русского происхождения Фебюс Арон Теодор Левен[22]впервые установил, что в состав нуклеиновой кислоты входит рибоза. До того считалось, что рибоза в природе не встречается. В 1901 году ее синтезировал Эмиль Фишер (первооткрыватель структуры пептидов), но считалось, что это соединение — не более чем научный курьез, не имеющий практического значения. Так гадкие утята превращаются впоследствии в прекрасных лебедей — и в науке, и в жизни...
Рибоза отличается от глюкозы, фруктозы и галактозы тем, что в ней не шесть, а пять атомов углерода. Эта пятизвенная углеродная цепочка стремится принять кольцевидную форму с помощью атома кислорода одной из гидроксильных групп. В результате получается кольцо из четырех атомов углерода и одного атома кислорода — можно считать, что это фурановое кольцо без двойных связей. На рис. 39 приведена структурная формула рибозы в двух видах — в полном и в «ломаном».

Рис. 39. Рибоза
Позже Левен обнаружил, что не все молекулы нуклеиновой кислоты содержат рибозу. В некоторых вместо нее имеется близкий к рибозе сахар, отличающийся лишь отсутствием одного из атомов кислорода. Соответственно химическое название этого сахара — дезоксирибонуклеиновая кислота, а строение его показано на рис. 40. Дезоксирибозу, как и рибозу, Фишер синтезировал за несколько лет до того, как ее обнаружили в природе.
По принципу наличия одного из этих двух сахаров нуклеиновые кислоты разделили на два подвида: рибонуклеиновая кислота и дезоксирибонуклеиновая кислота. Поскольку эти названия стали упоминаться все чаще, а биохимики любят большие и сложные названия не больше нас с вами, очень быстро в широкое употребление вошли сокращенные наименования этих веществ — рибонуклеиновая кислота стала РНК, а дезоксирибонуклеиновая — ДНК. Теперь полными именами их никто уже давно не называет.

Рис. 40. Дезоксирибоза
Кроме рибозы и дезоксирибозы, никаких других сахаров в нуклеиновых кислотах больше не обнаружилось, и за 1950-е годы биохимики пришли к выводу, что и не обнаружится. Соответственно единственно встречающимися в нуклеиновой кислоте сахарами являются рибоза и дезоксирибоза. Более того, не было обнаружено ни одной нуклеиновой кислоты, в состав которой входили бы и рибоза, и дезоксирибоза. Либо-либо.
Нуклеиновая кислота того или другого вида встречается в определенных участках клетки. ДНК находится исключительно в ядре — если быть совершенно точным, то именно в хромосомах. РНК тоже можно найти в ядре, но все же большая часть ее сосредоточена вне ядра, в цитоплазме. Насколько нам известно, все клетки содержат как РНК, так и ДНК.
Что же касается вирусов, то наиболее сложно организованные из них имеют в своем составе и ДНК, и РНК, большинство же — только ДНК. Впрочем, простейшие из вирусов, такие как вирус табачной мозаики, содержат только РНК.
ПУРИНЫ И ПИРИМИДИНЫ
Кроме фосфатных групп и сахаров, в нуклеиновых кислотах были обнаружены также соединения атомов, сгруппированных вокруг азотсодержащих колец. Этот факт установил Коссель в своих экспериментах в 1880-х годах и позже (затем он будет работать с протаминами). Все выделенные азотсодержащие соединения оказались выстроенными вокруг одной из двух кольцевых систем — пуринового кольца или пиримидинового кольца. Оба этих кольца мы уже приводили в данной книге на рис. 15. Поэтому азотсодержащие соединения, выделенные из нуклеиновой кислоты, объединяют в две группы — пуринов и пиримидинов[23].
Из нуклеиновых кислот в больших количествах были выделены два пурина и три пиримидина. Пурины — это аденин и гуанин, а пиримидины — цитозин, тимин и урацил. Все пять веществ представлены на рис. 41 как в полном, так и в «ломаном» отображении. Три из этих пяти соединений — аденин, гуанин и цитозин — присутствуют как в ДНК, так и в РНК. Тимин встречается только в ДНК, а урацил — только в РНК. Между собой эти два последних пиримидина различаются не очень — единственную разницу составляет тот факт, что у тимина есть метиловая группа, а у урацила — нет. Таким образом, в «ломаном» отображении на молекуле тимина будет иметься черточка, а на молекуле урацила — нет. Что касается генетического кода, то, забегая немного вперед, можно сказать, что тимин ДНК — это аналог урацила в РНК.

Рис. 41. Пурины и пиримидины
И еще по поводу формул: в некоторых органических соединениях атом водорода может достаточно свободно перемещаться, связываясь то с одним, то с другим атомом. Такое случается при наличии двойных связей, и переключение атома водорода подразумевает также и переключение двойных связей.
Так, например, в урациле атомы водорода гидроксильных групп с легкостью переключаются на ближайшие атомы азота в кольце. На самом деле они даже более склонны крепиться к атомам азота, чем к своим гидроксильным группам. Такое поведение атома водорода называется таутомерией. Формула таутомерного урацила приведена на рис. 42. Если сравнить его с рис. 41, где нарисована стандартная формула урацила, то видно, что единственной разницей, по крайней мере в «ломаном» отображении, будет изменение расположения двойных связей.

Рис. 42. Таутомерный урацил
Сам феномен таутомерии нас в дальнейшем интересовать не будет. Единственная причина, по которой мы о нем вообще упоминаем, — то обстоятельство, что время от времени нам надо будет нарисовать формулу какого-нибудь вещества вроде урацила в его таутомерной разновидности. Если бы сейчас мы об этом не упомянули, вас могла бы сбить с толку неожиданная разница в распределении двойных связей от формулы к формуле.
В крайне небольшом количестве образцов нуклеиновой кислоты была обнаружена пара мелких пиримидинов, представляющих собой видоизмененный цитозин. Мы не будем обращать на это внимание, поскольку в отношении генетического кода все они ведут себя так же, как цитозин. Так что в дальнейшем нам понадобится знать только те два пурина и три пиримидина, которые были здесь перечислены.
СКЛАДЫВАЕМ ВОЕДИНО
Итак, теперь перед нами полный список. Фосфатная группа, рибоза, дезоксирибоза, два пурина и три пиримидина — вот и все составляющие нуклеиновой кислоты. Вот наши восемь «слов», по аналогии с двадцатью двумя «словами», составляющими белок.
Это обескураживает — казалось бы, вещества, несущие генетический код, должны быть, как минимум, не менее сложными, чем белки. На самом же деле все еще проще. Из этих восьми «слов» в ДНК не используется рибоза и урацил, а в РНК — дезоксирибоза и тимин. Так что каждая из двух разновидностей нуклеиновой кислоты состоит только из шести «слов».
Но что удерживает эти «слова» вместе? Левен, первый, кто выделил из нуклеиновых кислот рибозу и дезоксирибозу, разрешил и эту проблему. Он разложил нуклеиновую кислоту на крупные части, каждая из которых содержала по нескольку базовых соединений. Изучая эти крупные составляющие, он описал их строение. А в начале 1950-х годов английский химик сэр Александр Тодд сумел синтезировать вещества, представленные в формулах Левена, и выяснилось, что они действительно обладают свойствами веществ, выделенных из нуклеиновой кислоты. Только после этого предположения Левена были безоговорочно приняты всеми биохимиками[24].
Левен установил, что в рибозной или дезоксирибозной составляющей каждой молекулы нуклеиновой кислоты имеется фосфатная группа, с одной стороны, и пурин или пиримидин — с другой. Эти соединения получили название нуклеотиды.
В составе РНК все нуклеотиды конечно же содержат, разумеется, рибозную группу, а кроме нее — одно из четырех соединений: аденин, гуанин, цитозин или урацил. Таким образом, могут иметься четыре различных нуклеотида: адениловая кислота, гуаниловая кислота, цитидиловая кислота и уридиловая кислота. И опять же — кислотные свойства каждой из них придает именно присутствие фосфатной группы, а из названия каждого нуклеотида ясно следует, какой именно пурин или пиримидин в нем присутствует.
Поскольку эти нуклеотиды играют крайне важную роль в генетическом коде, на рис. 43 я привожу формулы всех четырех, но только в «ломаном» виде.
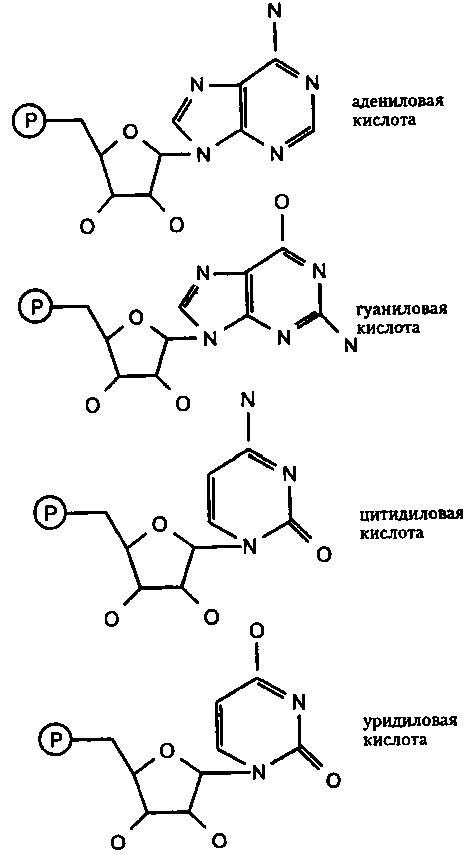
Рис. 43. Нуклеотиды рибозы
Нуклеотиды ДНК отличаются наличием дезоксирибозы вместо рибозы. Поэтому мы имеем в этом случае дезоксиадениловую кислоту, дезоксигуаниловую кислоту и дезоксицитидиловую кислоту. Дезоксиуридиловой кислоты в ДНК не бывает — поскольку вместо урацила в ней присутствует тимин, то и кислота получается дезокситимидиловая, как показано на рис. 44. Она отличается от уридиловой кислоты, как видите, отсутствием гидроксильной группы в сахаре. Дезоксиадениловая кислота отличается от адениловой, как и дезоксигуаниловая от гуаниловой, а дезоксицитидиловая — от цитидиловой.

Рис. 44. Дезокситимидиловая кислота
Все эти различия в нуклеотидах крайне важны для биохимии организма. Существуют нуклеотиды, подобные представленным на рис. 43, но имеющие по две или даже три объединенные фосфатные группы вместо одной. Эти соединения являются ключевыми в обеспечении хранения и высвобождения энергии; самым известным из них является аденозинтрифосфорная кислота, обычно обозначаемая сокращением АТФ. Молекула ее похожа на молекулу адениловой кислоты, но, в отличие от нее, в АТФ, как явствует из названия, имеются три фосфатные группы, а не одна.
Существуют также нуклеотидоподобные соединения, «кооперирующиеся» в совместных действиях с некоторыми ферментами и именуемые поэтому коферментами. В них иногда рибозу сменяет глюкоза или другие углеводы, а пурины или пиримидины иные азотсодержащие кольца.
Однако нас будут волновать только нуклеотиды нуклеиновой кислоты, а их в одной молекуле может содержаться только четыре разновидности.
Теперь мы можем задаться вопросом: каким же образом нуклеотиды собираются воедино для формирования нуклеиновой кислоты? Ответ на этот вопрос тоже был сформулирован Левеном и подтвержден Тоддом.
Ключ к разгадке — фосфатная группа. В отдельных нуклеотидах, как правило, имеется первичный фосфат с одной связью, но вместо него может быть и вторичный фосфат, вторая валентность которого связана вторым нуклеотидом. Таким образом, с помощью вторичных фосфатов можно связать целое множество нуклеотидов — на рис. 45 приведена в качестве примера подобная группа уридиловых кислот.
Представленные на рис. 45 объединенные нуклеотиды составляют полинуклеотидную цепочку. Если полинуклеотид состоит из рибозных нуклеотидов (как на рис. 45), то у каждой сахарной группы в цепочке имеется "торчащая в сторону" гидроксильная группа. Ее обозначают так: — О, отходящий от каждого сахарного кольца.
В тех полинуклеотидах, которые состоят из дезоксирибозных нуклеотидов, такой свободной гидроксильной группы нет (сравните рис. 44 и 43). Следовательно, РНК состоит из полинуклеотидной цепочки, у сахарной составляющей которой имеется гидроксильная группа, а ДНК — из полинуклеотидной цепочки, где сахар лишен гидроксильных групп.
Полинуклеотидная цепочка в определенном смысле похожа на полипептидную цепочку белков.

Рис. 45. Полинуклеотидная цепочка
Полипептидная цепочка состоит из «полиглициновой основы», которая тянется по всей длине молекулы, объединяя ее, и от которой во все стороны отходят различные радикалы, создающие необходимое разнообразие белковых молекул. То же самое можно сказать и о полинуклеотидной структуре, имеющей «сахаро-фосфатную основу», тянущуюся по всей длине молекулы, и отходящие от нее различные пурины и пиримидины. Схема сравнения приводится на рис. 46.

Рис. 46. Сравнительное строение белка и нуклеиновой кислоты
Молекулы белка различаются только радикалами, а молекулы нуклеиновой кислоты — только пуринами и пиримидинами.
Но, скажете вы, к полиглициновой основе могут крепиться двадцать два различных радикала, а к сахаро-фосфатной основе — только четыре различных элемента, пурина или пиримидина. Как же может нуклеиновая кислота, имея всего четыре «слова» в своем кодирующем «языке», нести достаточно информации, чтобы на ее основе можно было построить молекулу, «язык» которой будет обладать двадцатью двумя «словами»?
Всему свое время — до этого вопроса мы тоже обязательно дойдем и найдем на него ответ, но лишь после того, как рассмотрим поближе саму молекулу нуклеиновой кислоты.
Глава 8
ОТ ЦЕПОЧКИ К СПИРАЛИ
ДЛИНА ЦЕПОЧКИ
Теперь, когда нам известно, что именно связывает воедино нуклеотиды в молекуле нуклеиновой кислоты, можно задаться вопросом о том, сколько же нуклеотидов может объединять одна молекула.
До 1940-х годов большинство биохимиков над этим просто не задумывались. Прежде всего, само собой подразумевалось, что молекула нуклеиновой кислоты должна быть сравнительно небольшой. В пользу такого воззрения свидетельствовал тот факт, что она связывалась с белком, а ведь казалось естественным, что белок должен занимать доминирующее положение в любом соединении, в котором он присутствует.
Взять, например, гемоглобин. Кроме своих 574 аминокислот, он включает в себя еще и четыре гемовые группы, каждая из которых примерно в пять раз больше средней аминокислоты; соответственно, все четыре в сумме составляют где-то около 3% всей молекулы гемоглобина. Группы, подобные гемовым, называют простетическими, от греческого «prosthetikos» — «прибавляющий», от которого происходит и слово «протез».
Гем — это «рабочая часть» гемоглобина. В середине каждой гемовой группы содержится атом железа, к которому слабыми связями присоединяются молекулы кислорода, что и делает гемоглобин носителем кислорода в организме. Однако функционирование гемовой группы определяет на самом деле именно белковая часть. В организме есть ферменты, которые тоже содержат гемовые группы, например — каталаза, пероксидаза, различные цитохромы. Но выполнять функцию гемоглобина ни один из них не способен. И функции всех перечисленных белков тоже различны, и различия эти определяются именно различиями в белковой части молекулы.
Существуют и другие виды сложных белков, имеющих другие простетические группы. Например, гликопротеины, простетические группы которых представляют собой модификации сахаров. И каждый раз простетическая группа оказывается небольшим дополнением к самому белку, играя довольно незначительную роль. Так что вполне естественно было предположить, что и нуклеиновые кислоты тоже являются сравнительно небольшими соединениями, выполняющими некие второстепенные функции в составе общей молекулы.
Наблюдения самого Левена тоже, казалось, подтверждали это «само собой разумеющееся» предположение. Он выделил из нуклеопротеинов вещества, которые по дальнейшем рассмотрении оказались нуклеотидными цепочками длиной в среднем по четыре нуклеотида. Соответственно, вещества эти были названы тетрануклеотидами. Левен решил, что они-то и представляют собой простетические группы нуклеопротеинов. Из такого решения вполне логично проистекало предположение о том, что составлять тетрануклеотид должны все четыре различных нуклеотида.
К сожалению, на основе имевшейся у Левена информации невозможно было составить истинной картины. Его методика отделения нуклеиновой кислоты от белка подразумевала использование кислот и щелочных металлов. Эти вещества помогали высвободить нуклеиновую кислоту, но нуклеотидную цепочку они при этом дробили на мелкие части. Именно эти части и мог единственно наблюдать Левен.
Со временем другие биохимики разработали более щадящие способы выделения нуклеиновой кислоты, и им удалось добиться уже других результатов. Они выделяли нуклеиновые кислоты, состоявшие из гораздо более длинных нуклеотидных цепочек. Мало-помалу тетрануклеотидная теория строения нуклеиновой кислоты зашаталась под давлением новых фактов. В течение 1940-х годов химики выделяли все более и более длинные нуклеиновые кислоты, а к началу 1950-х были получены образцы РНК с молекулами, состоящими из тысячи нуклеотидов, и образцы ДНК с молекулами из двадцати тысяч нуклеотидов.
Но скорее всего, эти последние цифры являются завышенными. Вполне возможно, в процессе химической обработки для выделения случалось так, что несколько разных молекул нуклеиновой кислоты объединялись в одну с помощью слабых связей и таким образом получались «супермолекулы», в природе не существующие.
На сегодняшний день принято считать, что отдельный ген состоит из молекулы нуклеиновой кислоты, представляющей собой цепочку из 200-2000 нуклеотидов.
РАЗНОРОДНОСТЬ ЦЕПОЧКИ
Даже после того как стало известно, что молекула нуклеиновой кислоты может быть не меньше или даже больше, чем молекула белка (нуклеиновая кислота, состоящая всего из 200 нуклеотидов, имеет размер не меньший, чем молекула гемоглобина), тетрануклеотидная теория не сразу отмерла, а сначала просто приобрела модифицированный характер. С тем, что молекула нуклеиновой кислоты может представлять собой нечто большее, чем цепочку из четырех различных нуклеотидов, все согласились, но было выдвинуто предположение, что в таком случае эта молекула — просто многократное повторение все той же четырехзвенной цепочки.
Если бы подобным образом «уточненная» тетрануклеотидная теория была верна, то конечно же нуклеиновые кислоты никак не могли бы быть носителями генетического кода. Как молекула крахмала представляет собой лишь бесконечный повтор «глюкоза-глюкоза-глюкоза...», так и нуклеиновая кислота бесконечно повторяла бы в своем составе «тетрануклеотид-тетрануклеотид-тетрануклеотид...». В этом отношении совершенно не важно, что молекула тетрануклеотида в 7,5 раза крупнее молекулы глюкозы. Фраза, выглядящая как «непобедимость-непобедимость-непобедимость...», несет в себе не больше смысла, чем фраза «вот-вот-вот-вот...», несмотря на то что повторяющееся в нем слово длиннее и более значимо.
Однако после того, как в 1944 году были обнародованы эксперименты Эйвери, Мак-Леода и Мак-Карти (см. главу 6), биохимики с большой неохотой стали приходить к выводу, что тетранук-леотидную теорию не в силах спасти уже никакие модификации. Нуклеиновая кислота все-таки выступает носителем генетической информации, а по тетрануклеотидной модели этого получаться не могло. Более того, в ходе изучения трансформаций бактерий выяснилось, что нуклеиновые кислоты крайне разнообразны, что определенная нуклеиновая кислота вызывает определенные и только ей свойственные трансформации. Этого в рамках тетрануклеотидной теории объяснить было невозможно.
Теперь нуклеиновые кислоты подверглись более тщательному и интенсивному изучению.
К счастью, в том же 1944 году, когда Эйвери, Мак-Леод и Мак-Карти полностью перевернули взгляды на нуклеиновые кислоты, Мартин и Синг разработали технологию бумажной хроматографии. Изначально эта технология была разработана для аминокислот, но адаптировать ее под пурины и пиримидины оказалось несложно[25]. Соответственно нарисовалось четкое направление работ: разложить нуклеиновую кислоту, разделить пурины и пиримидины, проанализировать пурино-пиримидиновую смесь с помощью бумажной хроматографии и проверить, все ли вещества в ней представлены в равных количествах.
Если в равных, то тетрануклеотидная теория, возможно, и верна. Согласно тетрануклеотидной теории, пурины и пиримидины должны распределяться по схеме 1-2-3-4-1 -2-3-4-1 -2-3-4-..., соответственно количество их будет одинаково. Однако и при одинаковом количестве самих элементов все еще остается возможность их неравномерного распределения.
С другой стороны, если бы анализ пуриново-пиримидиновой связи показал, что отдельные составляющие представлены в ней в неравных количествах, то это однозначно свидетельствовало бы о том, что тетрануклеотидная теория неверна.
Так и оказалось. Одним из самых дотошных исследователей этой проблемы был Эрвин Чаргафф. К 1947 году он добился результатов, четко свидетельствовавших не только о том, что количество пуринов и пиримидинов в нуклеиновых кислотах различно, но и что от одной молекулы нуклеиновой кислоты к другой соотношение различных нуклеотидов тоже меняется. Так закончилась история тетрануклеотидной теории.
В начале 1950-х годов Чаргафф смог еще убедительнее показать, что различные нуклеотиды, казалось бы, случайным образом разбросаны по нуклеиновой кислоте. В таком случае количество возможных вариаций полинуклеотидной цепочки представлялось огромным — хотя и не столь огромным, как количество вариаций полипептидной цепочки аналогичной длины, ведь полипептидная цепь располагает 22 элементами для распределения, а полинуклеотидная — только четырьмя.
К примеру, количество возможных вариантов полипептидной цепочки длиной в 20 аминокислот составляет чуть больше 2 400 000 000 000 000 000 (примерно два с половиной квинтиллиона), а полинуклеотидная цепочка длиной в 20 нуклеотидов, по пять каждой из четырех их разновидностей, может быть представлена лишь чуть более чем 1 100 000 000 комбинаций.
Иными словами, вариативность полипептидной цепочки более чем в два миллиарда раз превышает вариативность полинуклеотидной цепочки аналогичной длины.
Но ведь никто и не обязывает полинуклеотидные цепочки иметь не больше звеньев, чем имеют цепочки, образующие белки! Если взять некую полипептидную цепочку, содержащую определенное количество аминокислот, и полинуклеотидную цепочку, содержащую в два раза больше нуклеотидов, то их вариативность будет уже примерно одинаковой. Бедность знакового набора нуклеотидов (4 знака против 22 у белков) вполне может компенсироваться увеличением длины цепочки!
Вообще, складывается впечатление, что средняя молекула нуклеиновой кислоты не в два, а в пять раз длиннее средней белковой молекулы. Так что в итоге вариативность оказывается выше не у белка, а у нуклеиновой кислоты.
К началу 1950-х
|
|
|
