 |


Регуляция депонирования и мобилизации гликогена
|
|
|
|
Синтез гликогена
Гликоген синтезируется практически во всех тканях, особенно активно в печени и скелетных мышцах. Синтез происходит в период пищеварения, спустя 1-2 ч после приема углеводной пищи. Синтез гликогена требует энергии. При включении одного мономера в полисахаридную цепь протекают 2 реакции, сопряженные с расходованием АТФ и УТФ.
 |
Перенос глюкозильных групп от УДФ-глюкозы на нередуцирующий конец разветвленной молекулы гликогена катализируется ферментом гликогенсинтазой. При этом образуется новая a-1,4-гликозидная связь. Образование a-1,6-связей, находящихся в точках ветвления цепей гликогена, гликогенсинтаза не способна. Эти связи образует специальный «ветвящий» фермент путем переноса коротких фрагментов (из 6-7 остатков глюкозы) с одного участка гликогена на другой с образованием a-1,6-гликозидных связей. Таким образом, наращивание молекулы гликогена осуществляется путем чередования действия этих двух ферментов.
Регуляция депонирования и мобилизации гликогена
Переключение процессов синтеза и мобилизации гликогена в печени происходит при переходе состояния пищеварения в постабсорбтивный период или состояния покоя на режим мышечной работы. В переключении этих метаболических путей в печени участвуют гормоны инсулин, глюкагон и адреналин, а в мышцах – инсулин и адреналин.
Влияние этих гормонов на синтез и распад гликогена реализуется на уровне ключевых ферментов - гликогенсинтазы и гликогенфосфорилазы – и осуществляется путем их фосфорилирования и дефосфорилирования.
Первичным сигналом для синтеза инсулина и глюкагона является изменение концентрации глюкозы в крови. Инсулин и глюкагон постоянно присутствуют в крови, но при переходе из абсорбтивного состояния в постабсорбтивное изменяется их относительная концентрация – инсулин-глюкагоновый индекс (ИГИ). Т.о., главным переключающим фактором в печени является инсулин ИГИ.
|
|
|
В постабсорбтивном периоде ИГИ снижается и решающим фактором является влияние глюкагона, который стимулирует распад гликогена в печени. Механизм действия глюкагона включает каскад реакций, приводящий к активации гликогенфосфорилазы:
аденилатциклаза (неакт)
гормон® ¯ АТФ
аденилатциклаза (акт) ® ¯ ПК А (неакт)
цАМФ® ¯ киназа фосфорилазы (неакт)
ПК А (акт)® ¯ фосфорилаза В
киназа фосфорилазы (акт)® ¯ гликоген
фосфорилаза А→ ↓
глюкозо-1-Ф
В период пищеварения преобладающим является влияние инсулина, т.к. ИГИ в этом случае повышается. Под влиянием инсулина происходит:
а) стимуляция транспорта глюкозы в клетки мышечной ткани.
б) изменение количества некоторых ферментов путем индукции и репрессии их синтеза. Например, инсулин индуцирует синтез глюкокиназы, ускоряя тем самым фосфорилирование глюкозы в печени.
в) изменение активности ферментов путем фосфорилирования и дефосфорилирования. Инсулин снижает содержание цАМФ в клетке (а значит и цАМФ-зависимое фосфорилирование) путем активации фосфодиэстеразы – фермента, гидролизующего цАМФ, с одной стороны. С другой стороны, он активирует фосфатазу гликогенсинтазы. Последняя дефосфорилируется и переходит в активное состояние.
Пентозофосфатный путь превращений глюкозы
ПФП является альтернативным путем окисления глюкозы. Он не приводит к синтезу АТФ, но поставляет клеткам кофермент НАДФН·Н+, а также обеспечивает клетки рибозой. НАДФН·Н+, использующийся как донор водорода в реакциях восстановления, участвует в реакциях биосинтеза жирных кислот и холестерина, а также в реакциях гидроксилирования, играющих важную роль в функционировании микросомальной цепи обезвреживания чужеродных веществ. Рибоза участвует в синтезе нуклеотидов и нуклеиновых кислот, а также аминокислоты гистидина.
|
|
|
ПФП может функционировать в печени, жировой ткани, молочной железе, коре надпочечников, эритроцитах и органах, где активно протекают восстановительные синтезы, например синтез жирных кислот. Все ферменты ПФП локализованы в цитозоле.
Химизм
1 реакция: дегидрирование глюкозо-6-фосфата
Реакцию катализирует глюкозо-6-фосфатдегидрогеназа, ключевой фермент пентозофосфатного цикла. Образовавшийся в ходе реакции 6-фосфоглюконолактон - соединение нестабильное, которое спонтанно, либо под действием специфической лактоназы гидролизуется.
2 реакция: гидролиз 6-фосфоглюконолактона с образованием 6-фосфоглюконата. Равновесие суммарной реакции сильно смещено в сторону образования НАДФ·Н+.
3 реакция: дегидрирование и декарбоксилирование 6-фосфоглюконата с образованием рибулозо-5-фосфата.
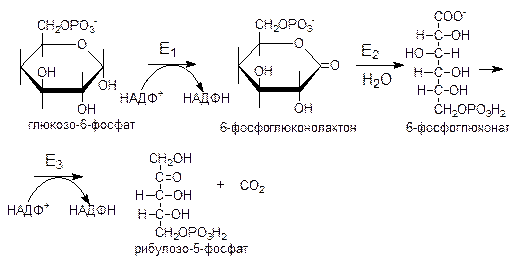 |
Превращение глюкозо-6-фосфата до рибулозо-5-фосфата принято называть окислительной фазой пентозофосфатного цикла. Фаза от рибулозо-5-фосфата до образования вновь глюкозо-6-фосфата называется неокислительной или анаэробной фазой этого цикла.
4 реакция: изомеризация пентозофосфатов. Рибулозо-5-фосфат под действием ферментов рибозофосфатизомеразы и рибулозо-5-фосфат-3-эпимеразы может обратимо изомеризоваться в другие пентозы: рибозо-5-фосфат, ксилулозо-5-фосфат.
 |
В некоторых случаях ПФП на этом заканчивается. И тогда суммарное уравнение реакции выглядит так:
глюкозо-6-фосфат + Н2О + 2НАДФ+ ® рибозо-5-фосфат + 2НАДФН·Н+ +СО2
Однако многие клетки нуждаются в большем количестве НАДФН·Н+ (для восстановительных синтезов), чем требуется рибозо-5-фосфата (для включения в нуклеотиды). В таких случаях рибозо-5-фосфат превращается в глицеральдегид-3-фосфат и фруктозо-6-фосфат под действием двух ферментов: транскетолазы и трансальдолазы. Эти ферменты создают обратимую связь между ПФП и гликолизом, катализируя следующие реакции:
5) ксилулозо-5-фосфат + рибозо-5-фосфат ® седогептулозо-7-фосфат + глицеральдегид-3-фосфат
|
|
|
6) седогептулозо-7-фосфат + глицеральдегид-3-фосфат ® эритрозо-4-фосфат + фруктозо-6-фосфат*
7) ксилулозо-5-фосфат + эритрозо-4-фосфат ® фруктозо-6-фосфат* + глицеральдегид-3-фосфат*
Суммируя эти реакции, получаем: 2 ксилулозо-5-фосфат + рибозо-5-фосфат ® 2 фруктозо-6-фосфат + глицеральдегид-3-фосфат
Итак, избыток рибозо-5-фосфата, образованный в ПФП, может превращаться в метаболиты гликолиза.
Метаболизм фруктозы
Фруктоза, поступающая в организм человека, образуется либо при гидролизе сахарозы, либо поступает в составе растений, являясь их основным сахаром. У здорового человека фруктоза может превращаться по двум путям: 1) под действием гексокиназы (в печени, мышцах, почках, жировой ткани) образуется фруктозо-6-фосфат. Однако, сродство этого фермента к фруктозе гораздо ниже, чем к глюкозе, поэтому в присутствии глюкозы реакция фосфорилирования фруктозы протекает с низкой скоростью; 2) под действием фруктокиназы (в печени, почках и кишечнике) образуется фруктозо-1-фосфат. Фермент обладает высоким сродством к своему субстрату и в присутствии глюкозы становится основным путем метаболизма фруктозы.
 |
Фруктоза включается в гликолиз на стадии триозофосфатов.
Е1 – фруктокиназа
Е2 – фруктозо-1-фосфатальдолаза
Е3 - триозофосфатизомераза
Расщепление фруктозы по пути гликолиза происходит быстрее, чем глюкозы, т.к. минуется стадия, катализируемая фосфофруктокиназой-I.
Метаболизм фруктозы в сперматозоидах
Фруктоза является основным источником энергии для сперматозоидов. Она образуется в семенных пузырьках из глюкозы в результате 2-х последовательных реакций:
1)D-глюкозы + НАДФН·Н+ ® D-сорбитол + НАДФ+
реакция катализируется сорбитолдегидрогеназой.
2) D-сорбитол + НАД+ ® D-фруктоза + НАДН·Н+
реакция катализируется сорбитолдегидрогеназой.
Концентрация фруктозы в семенной жидкости может достигать 10 мМ.
Метаболизм галактозы
Галактоза, поступающая в организм человека, образуется при гидролизе лактозы (молочного сахара). Галактоза включается в метаболизм путем превращения в глюкозо-1-фосфат.
|
|
|
 |
Образовавшийся глюкозо-1-фосфат после превращения в глюкозо-6-фосфат подключается к основным путям метаболизма глюкозы.
|
|
|
