 |


Синтез незаменимых аминокислот
|
|
|
|
Высшие растения способны синтезировать все необходимые для белкового синтеза аминокислоты и могут использовать для этого соответствующие a-кетокислоты, а также аммиак или нитраты в качестве источника азота. Организм животных и человека не синтезирует все необходимые аминокислоты. Не синтезируются только 10 из 20 необходимых, или незаменимых аминокислот: валин, лейцин, изолейцин, треонин, метионин, фенилаланин, триптофан, лизин, аргинин, гистидин.
Незаменимость этих аминокислот для роста и развития организма животных и человека объясняется отсутствием соответствующих a-кетокислот, из которых в процессе аминирования они могли бы синтезироватся.
Биосинтез каждой незаменимой аминокислоты имеет специфические особенности; он гораздо сложнее, чем биосинтез заменимых аминокислот.
Поскольку синтез каждой из незаменимых аминокислот имеет свою особенность, ограничимся лишь некоторыми общими положениями. Так, три незаменимых аминокислоты лизин, треонин и метионин в растениях и микроорганизмах синтезируются из аспарагиновой кислоты.
Изолейцин образуется у бактерий из незаменимой аминокислоты треонина. Многостадийный синтез у растений фенилаланина осуществляется из эритрозо-4-фосфата и фосфопирувата; на предпоследних стадиях осуществляется перенос аминогруппы от глутаминовой кислоты. Биосинтез гистидина – незаменимой аминокислоты для детей – полностью изучен у бактерий и грибов. Завершающей стадией является реакция трансаминирования, роль донора аминогруппы выполняет также глутаминовая кислота. Путь биосинтеза гистидина в высших растениях не изучен. Синтезированные аминокислоты – заменимые и незаменимые используются для биосинтеза белка.
|
|
|
Биосинтез белка
Выявление механизма биосинтеза белка – одна из самых важных и интересных проблем современной биохимии. Приблизительные подсчеты показывают, что одна живая клетка содержит несколько тысяч разных белков, организм же в целом должен синтезировать десятки тысяч индивидуальных белковых молекул.
Каким же образом в клетке образуется такое большое количество разнообразных белков из небольшого набора аминокислот, причем именно в то время и в таком количестве, в каком требуется? И как специфические свойства белков передаются из поколения в поколение? Такие вопросы волнуют ученых во всем мире. Механизм сложных жизненных процессов, лежащих в основе передачи наследственности, начали расшифровывать лишь в последние 60 лет.
Процесс биосинтеза белков оказался универсальным для всех живых существ на Земле – от простейшей бактериальной клетки до высших растений, животных и человека. Синтез белка в клетке основывается на двух фундаментальных принципах, характерных для живых систем, отличающих биологические системы от неживой природы, – матричный принцип и принцип комплементарности.
Матричный принцип состоит в том, что взаимодействие происходит не между молекулами, находящимися в системе в хаотическом движении, а между пространственно организованными фиксированными молекулами и системами.
Одно из этих веществ обязательно представляет собой полимер, тогда как другое может быть как полимером, так и мономером. Матричный синтез является основным во всех тех случаях, когда необходимо обеспечить заранее заданную последовательность мономеров во вновь синтезированном биополимере.
Принцип матричного синтеза реализуется через принцип комплементарности. Именно комплементарность позволяет матрице «выбрать необходимый мономер и установить его в нужном месте на матрице».
|
|
|
В результате кропотливой работы многих ученых была установлена в основном роль нуклеиновых кислот в биосинтезе белков и показана матричная роль РНК в этом процессе, что позволило Ф. Крику выработать положение о передаче генетической информации в клетке.
Последовательность матричных биосинтезов белка включает три основных этапа.
1) Репликацию ДНК – биосинтез копии ДНК с использованием в качестве матрицы уже существующие молекулы ДНК.
2) Транскрипцию – биосинтез РНК (любой тРНК, мРНК, рРНК) на матрице ДНК. Нуклеотидная последовательность в молекуле РНК комплементарна какому-то участку (гену) молекулы ДНК.
3) Трансляция – биосинтез полипептидных цепей белковых молекул, аминокислотная последовательность которых задается нуклеотидной последовательностью мРНК при участии тРНК и рРНК. В качестве матрицы используется мРНК.
Синтез ДНК – репликация
Человека всегда интересовал вопрос, каким образом происходит передача наследственных признаков.
И вот на рубеже 19 и 20 веков генетические и цитологические исследования привели к выводу, что ответственными за передачу признаков по наследству являются хромосомы. При этом оказалось, что определенный признак передается с определенным участком хромосомы – геном.
Всему набору признаков организма (голубые глаза, темные волосы, прямой нос и т.д.) соответствует набор генов всех хромосом – генотип.
Поскольку хромосомы содержат белок и ДНК, возник вопрос, какое из веществ участвует в передаче наследственных признаков. В 40-50-е годы ХХ века появилось много экспериментальных данных на то, что передача наследственной информации осуществляется молекулами ДНК.
Общее содержание ДНК в одной клетке постоянно, количество ее в процессе жизненного цикла клетки практически не меняется.
Но в период деления клетки оно резко возрастает, и, как показали исследования, концентрация ДНК увеличивается ровно в два раза. После клеточного деления содержание ДНК в дочерних клетках такое же, каким оно было и в материнской клетке.
Каким же образом происходит удвоение, или как часто говорят, репликация молекул ДНК? Ответ на этот вопрос был получен на основании структурной модели строения ДНК, разработанной Уотсоном и Криком в 1953 г. Структура двойной спирали позволяла представить следующий механизм репликации ДНК.
|
|
|
При репликации молекул ДНК в период клеточного деления прежде всего разрываются водородные связи между цепями, двойная спираль раскручивается, цепи расходятся. Под действием соответствующих ферментов к каждой из одиночных цепей присоединяются новые нуклеотиды. Но так как сочетание нуклеотидов может быть только строго определенным, то на каждой образовавшейся цепи строится вторая комплементарная цепь, образуя новые молекулы ДНК прежнего состава.
Процесс удвоения ДНК представлен на схеме:

Реакцию синтеза ДНК можно представить такой схемой:
m(gАТФ + gТТФ) + n(gГТФ + gЦТФ) 
ДНК + (m +n) Н4Р2О7
Отметим важнейшие особенности реакции:
1. Субстратами служат трифосфаты дезоксирибонуклеозидов. В ходе реакции от каждого из них отщепляется пирофосфатный остаток; при этом выделяется энергия, которая затрачивается на включение каждого мономера в молекулу ДНК.
2. Реакция идет в присутствии готовой ДНК, выполняющей роль матрицы. Все вновь синтезируемые молекулы ДНК имеют первичную структуру, идентичную первичной структуре ДНК-матрицы.
3. Поскольку в молекуле ДНК нуклеотидные остатки образуют пары А-Т; и Г-Ц, то в реакции расходуются одинаковое количество дАТФ и дТТФ и одинаковое количества дГТФ и дЦТФ.
Так как в молекуле ДНК записана генетическая информация (т.е. какие белки должны синтезироваться в клетке), то процесс удвоения ДНК в материнской клетке и последующее распределение копий поровну между дочерними клетками есть ни что иное, как передача генетической информации от поколения к поколению. Такой способ репликации получил название полуконсервативного. Репликация происходит при участии сложного набора белков, образующих репликативный комплекс. В него, в частности, входят белки, раскручивающие спираль ДНК, в результате чего образуется репликативная вилка. Затем при участии ДНК-полимеразы (репликазы) образуются новые полинуклеотидные цепи. Синтез новых цепей всегда идет в направлении от 5¢- конца к 3¢- концу. Поэтому на одной из ветвей репликативной вилки новая цепь наращивается непрерывно по мере раскручивания ДНК- матрицы. На другой ветви по мере раскручивания ДНК образуются короткие фрагменты новой цепи – фрагменты Оказаки. Затем концы этих фрагментов соединяются между собой в результате действия ДНК- лигазы (рис 2).
|
|
|


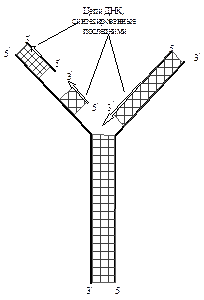
Рисунок 2 – Схема репликации ДНК
В данной схеме приведены лишь основные этапы репликации; в действительности этапов репликации значительно больше, в них участвуют специальные белки. В настоящее время установлено около десятка белков репликативного комплекса. Таким образом, благодаря принципу комплементарности полинуклеотидных цепей из одной исходной молекулы ДНК образуются две совершенно одинаковые, новые молекулы ДНК. Способность ДНК давать строго определенные, подобные себе, новые молекулы играют определяющую роль в явлениях наследственности и в передаче генетической информации от поколения к поколению.
Другими словами, информация, записанная в ДНК (в генотипе), обеспечивает образование фенотипических признаков (окраска цветка, цвет волос, форма крыльев и т.д.) организма, трансформируется в фенотип.
В 40-50-е годы это представление о связи между генотипом и фенотипом было экспериментально подтверждено на многих ферментах и других белках разных организмов; результаты нашли отражение в афористической формуле один ген – один белок. ДНК сосредоточена главным образом в ядре, синтез же белка происходит в другом месте (в рибосомах). Роль промежуточного переносчика информации от ДНК к месту синтеза белка выполняют рибонуклеиновые кислоты. Их роль в процессе биосинтеза белка различна.
Направление потока информации в клетке от генотипа к фенотипу представляют так:
ДНК ® РНК ® белки.
транскрипция трансляция
Иначе говоря, ДНК служит матрицей для синтеза всех РНК, а РНК – матрицей для синтеза белков. Это положение называют основным постулатом молекулярной биохимии.
Синтез РНК – транскрипция
Содержание РНК во всех клетках живого организма непостоянно. Оно может меняться в зависимости от вида, возраста, физиологического состояния организма и ткани. Обычно в молодых клетках содержание РНК больше, чем в старых. Кроме того, в клетках содержатся различные по своим функциям и молекулярной массе виды РНК – рибосомная, транспортная и матричная.
Все это обуславливает значительные трудности в изучении биосинтеза рибонуклеиновых кислот. Считают, что синтез РНК может происходить несколькими путями. Мы рассмотрим один из них, так называемый ДНК-зависимый синтез РНК. С точки зрения передачи наследственной информации в процессе биосинтеза белков, наибольшее значение имеет синтез молекул РНК на участке молекулы ДНК как на матрице.
|
|
|
Исходными веществами, необходимыми для биосинтеза, являются рибонуклеозидтрифосфаты (АТФ, ГТФ, ЦТФ, УТФ); синтез катализируется ферментом РНК-полимеразой, сходной по своему действию с ДНК-полимеразой. Схема биосинтеза следующая:
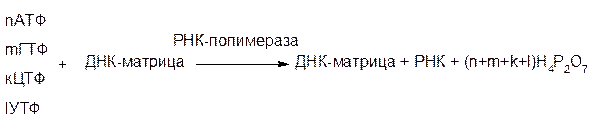
Как известно, ДНК представляет собой двойную спираль. Но при матричном синтезе РНК на участке молекулы ДНК «копируется» только одна из двух ее цепей, которую называют значащая. Она служит матрицей.
Все синтезированные молекулы РНК имеют структуру, комплементарную матрице (т.е участку – гену одной из цепи ДНК).
Прежде чем начнется образование цепи РНК, фермент РНК-полимераза присоединяется к матрице не в любом ее месте, а к определенному участку – который называется промотором. На этом участке молекулы ДНК есть последовательности нуклеотидов, узнаваемые РНК-полимеразой.
Связывание РНК-полимеразы с промотором приводит к локальному (местному) расхождению нуклеотидных цепей, в этом участке ДНК одна из цепей ДНК служит матрицей.
Наращивание молекулы РНК происходит в результате перемещения РНК-полимеразы вдоль ДНК путем присоединения очередного рибонуклеотида, комплементарного тому дезоксирибонуклеотиду ДНК, который в данный момент находится в области активного центра РНК-полимеразы.
В молекуле ДНК закодированы не только участки, с которых начинается синтез РНК, но и последовательности нуклеотидов, сигнализирующих о прекращении синтеза РНК. С этими участками, по-видимому, связываются определенные белки, останавливающие действие РНК-полимеразы. Таким образом, получаются отдельные молекулы РНК, каждая из которых содержит информацию одного гена. Синтез РНК происходит в направлении от 5¢- конца к 3¢- концу.
В клетках высших организмов обнаружено три типа ДНК-зависимых РНК-полимераз. Они катализируют синтез рибосомной, матричной и транспортной РНК.
Все виды РНК образуются на ДНК-матрице в виде крупных молекул предшественников, которые затем подвергаются значительным изменениям –созреванию. При созревании, под действием ферментов, от них постепенно отщепляются лишние нуклеотиды, а у тРНК происходит метилирование и восстановление части оснований.
Биологический код
Биосинтез белков, или трансляция, отличается от биосинтеза ДНК и РНК (репликации ДНК и трансляции РНК) двумя принципиальными особенностями: между мРНК (матрицей) и пептидной цепью белка, которая на этой матрице синтезируется, нет комплементарного соответствия, т.е. структура рибонуклеотидов (мономеров матрицы) и аминокислот (мономеров белка) такова, что избирательные взаимодействия между ними, подобные образованию пар А…Т или Г…Ц, невозможны, нет соответствия между числом мономеров – нуклеотидов в матрице и в продукте реакции (в мРНК 4 разных нуклеотида, в белке 20 разных аминокислот);
Поэтому механизм использования матрицы при биосинтезе белка отличается от использования матриц при репликации и транскрипции.
Способ записи информации о первичной структуре белков в нуклеиновых кислотах получил название биологического кода (его также называют генетическим, нуклеотидным, аминокислотным кодом).
Один из первых вопросов, который возникает при выяснении структуры биологического кода, – это вопрос о кодовом числе; т.е. о числе нуклеотидных остатков, кодирующих включение в белок одной аминокислоты. Очевидно, что кодовое число не может быть равным единице, так как в этом случае с помощью четырех нуклеотидов можно было бы закодировать только четыре аминокислоты. При кодовом числе 2 количество разных нуклеотидных пар будет равно числу перестановок из четырех элементов по 2, т.е равно 42 = 16, что тоже недостаточно для кодирования всех аминокислот. Число разных троек нуклеотидов равно 43 = 64. Это в три с лишним раза превышает минимальное число необходимое для кодирования 20 аминокислот. Экспериментально доказано, что в биологическом коде кодовое число равно трем: тройку нуклеотидных остатков (триплет), кодирующих включение одной аминокислоты, называют кодоном.
Из 64 кодонов (триплетов), (табл.1) 61 используется для кодирования аминокислот, а три – УАА, УAГ и УГA – обозначают конец матрицы: на этих триплетах обрывается дальнейшее наращивание пептидной цепи – терминирующие кодоны. Каждый триплет кодирует только какую-нибудь одну аминокислоту. Это свойство кода называют специфичностью. С другой стороны, одна аминокислота может кодироваться двумя или большим числом (до шести) разных триплетов, т.е. код вырожденный, но первые два нуклеотида для данной аминокислоты всегда одинаковы. Непрерывность кода белкового синтеза состоит в том, что все входящие в его состав кодоны располагаются в мРНК, кодирующей биосинтез определенного белка, в строгом порядке один возле другого, не будучи разделенными иными моно- или олигонуклеотидными вставками. Неперекрывающийся характер кода заключается в том, что ни один из нуклеотидов одного кодона не является составной частью другого (соседнего) кодона.
К настоящему времени биологический код изучен у всех биологических объектов (от вирусов и бактерий до высших животных). Во всех случаях он оказался одинаковым. Эта универсальность кода свидетельствует о единстве происхождения всех форм жизни на Земле.
Таблица 1 – Биологический код
| У | Ц | A | Г | ||||
| Первая буква | У | 

| 
| 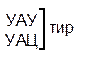

| 

| У Ц А Г | Третья буква |
| Ц | 
| 
| 

| 
|  У
Ц
А
Г
У
Ц
А
Г
| ||
| A | 
| 
| 
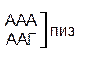
| 

| У Ц А Г | ||
| Г | 
| 
| 

| 
| У Ц А Г | ||
Генетический код, состоящий из 64 триплетов – оснований (кодонов) и соответствующих им аминокислот
3.4 Схема передачи генетической информации
от ДНК к белку
Допустим, что на определенном участке одной из спиралей молекулы ДНК, на которой синтезируется мРНК, мы имеем определенную последовательность нуклеотидов. Синтезированная на этом участке мРНК может иметь лишь строго определенную, комплементарную ДНК последовательность нуклеотидов (учитывая, что урацил РНК соответствует тимину ДНК). «Считывая» нуклеотидный код мРНК справа налево (от N-конца полипептида к С-концу), получим определенную последовательность аминокислот в данном полипептиде, который может синтезироваться под генетическим контролем этого участка молекулы ДНК.
Перенос информации от ДНК к белку можно изобразить в виде схемы:
| ДНК | Г А А | | | | | | ЦТ Т | А Ц Т | | | | | | Т Г A | Ц Г Г | | | | | | Г Ц Ц | A T T | | | | | | Т A A | – …Нетранскрибируемая цепь …Кодоны ДНК |
| мРНК | | | | Г А А | | | | А Ц У | | | | Ц Г Г | | | | A У У | …Кодоны мРНК |
| Белок | Глутами-новая кислота | Треонин | Аргинин | Изолейцин | …Аминокислотная последовательность |
|
|
|
