 |


в эпигенезе «ничто не возникает из ничего»
|
|
|
|
Как я уже заметил, эпигенез относится к эволюции, как разработка тавтологии к творческой мысли. В эмбриологии живого организма не только нет надобности в новой информации или в изменении плана, но большей частью эпигенез должен быть защищен от вторжения новой информации. Путь к этому – тот же, каким был всегда. Развитие зародыша должно следовать аксиомам и постулатам, изложенным в ДНК, или где-нибудь еще. На языке Главы 2, эволюция и обучение неизбежно дивергентны и непредсказуемы, а эпигенез должен быть конвергентным.
Отсюда следует, что в области эпигенеза случаи, когда требуется новая информация, редки и бросаются в глаза. Напротив, должны быть случаи, хотя и патологические, где недостаток или потеря информации приводит к серьезным расстройствам развития. В этом контексте, явления симметрии и асимметрии оказываются богатым источником примеров. Правила, которыми должен руководствоваться ранний эмбрион в этих отношениях, просты и формальны, так что их присутствие или отсутствие нельзя не заметить.
Наиболее известные примеры происходят из экспериментального изучения эмбриологии амфибий, и я рассмотрю здесь некоторые явления, связанные с симметрией яйца лягушки. То, что известно о лягушке, вероятно, справедливо для всех позвоночных.
Кажется, без информации из внешнего мира неоплодотворенное яйцо лягушки не имеет необходимой информации (т.е. необходимого различия) для достижения двусторонней симметрии. В яйце есть два дифференцированных полюса: животный полюс, где протоплазма преобладает над желтком, и растительный полюс, где преобладает желток. Но в нем нет дифференциации меридианов, или линий долготы. В этом смысле яйцо радиально симметрично.
|
|
|
Несомненно, дифференциация животного и растительного полюсов определяется положением яйца в фолликулярной* ткани, или плоскостью последнего клеточного делении при образовании гаметы¦; эта плоскость, в свою очередь, вероятно, определяется положением материнской клетки в фолликуле. Но это еще не все, что требуется.
Без некоторой дифференциации сторон или меридианов неоплодотворенного яйца оно не может «знать» или «решить», какова должна быть будущая средняя плоскость симметрии двусторонне симметричной лягушки. Эпигенез не может начаться, пока один из меридианов не будет выделен по сравнению с другими. К счастью, в этом случае мы знаем, как доставляется эта решающая информация. Она неизбежно приходит из внешнего мира: это точка входа сперматозоида. Как правило, сперматозоид входит в яйцо несколько ниже экватора, и меридиан, содержащий оба полюса и точку входа, определяет среднюю плоскость двусторонней симметрии лягушки. Первая сегментация яйца следует этому меридиану, и та сторона яйца, где входит сперматозоид, становится брюшной стороной лягушки.
Далее, известно, что необходимое сообщение об этом не содержится в ДНК или в других структурах сперматозоида. Той же цели достигает укол щетинки верблюжьего волоса. Вслед за таким уколом, яйцо будет сегментировать и продолжит свое развитие, превратившись во взрослую лягушку, которая будет прыгать и ловить мух. Конечно, она будет гаплоидной (т.е. у нее будет недоставать половины нормального комплекта хромосом). Она не будет размножаться, но в остальном будет во всех отношениях совершенна.
Для этого не нужен сперматозоид. Все, что нужно, – это указатель различия, и организм не особенно разборчив в отношении характера этого указателя. Без какого-нибудь указателя – не будет эмбриона. «Ничто не возникает из ничего».
Но это еще не все. Будущая лягушка – а в действительности уже очень молодой головастик – заметным образом асимметричен в анатомии своей эндодермы+. Подобно большинству позвоночных, лягушка довольно точно симметрична в своей эктодерме (кожа, мозг и глаза) и мезодерме (скелет и скелетные мускулы), но резко асимметрична в эндодермных структурах (кишки, печень, поджелудочная железа, и так далее). (В действительности, любое животное, у которого кишки складываются не в меридиональной плоскости, должно быть асимметрично в этом отношении. Если вы посмотрите на живот головастика, то вы отчетливо увидите, что кишки, ясно видимые через кожу, у него сложены в большую спираль).
|
|
|
Как можно ожидать, situs inversus (обращенная симметрия) у лягушек встречается, но крайне редко. Это явление известно у людей, и встречается в одном случае на миллион. Такие индивиды выглядят в точности, как все другие, но внутренне они обращены, так что правая сторона сердца связана с аортой, а левая с легкими, и так далее. Причины такого обращения неизвестны, но тот факт, что оно вообще случается, указывает, что нормальная асимметрия не определяется асимметрией молекул. Обращение какой-либо части такой химической асимметрии потребовало бы обращения всех ее частей, поскольку молекулы надлежащим образом подходят друг к другу. Обращение всей химии немыслимо, такой организм не мог бы выжить иначе как в обращенном мире.
Таким образом, остается проблема об источнике информации, определяющем асимметрию. Конечно, должна быть информация, указывающая яйцу правильную (статистически нормальную) асимметрию.
Насколько нам известно, после оплодотворения нет момента, когда могла бы быть доставлена такая информация. События следуют таким образом: сначала отделение от матери, затем оплодотворение; а после этого, в течение периода сегментации и раннего эмбрионального развития, яйцо защищено студенистой массой. Иными словами, яйцо несомненно должно содержать информацию, необходимую для определения асимметрии, уже до оплодотворения. В какой же форме эта информация должна существовать?
В Главе 3, при обсуждении природы объяснения, я заметил, что никакой словарь не может определить слова левое и правое. Это значит, что проблему не может решить никакая, как угодно устроенная цифровая система; информация должна быть прямо введена. Теперь у нас есть шанс обнаружить, как эта же проблема решается яйцом.
|
|
|
Как я полагаю, в принципе здесь может быть лишь решение одного рода (и я надеюсь, что кто-нибудь со сканирующим электронным микроскопом найдет подтверждение). Дело должно быть в том, что ответ заключен в яйце уже до оплодотворения, и следовательно в такой форме, что он определяет одну и ту же асимметрию независимо от того, какой меридиан отмечен входом сперматозоида. Отсюда следует, что любой меридиан, где бы он ни был проведен, должен быть асимметричен, и что все они должны быть асимметричны в одном и том же смысле.
Это требование проще всего удовлетворяется некоторой спиралью неколичественных или векторных отношений. Такая спираль должна пересекать все меридианы наискось, отмечая для каждого из них одну и ту же разницу между востоком и западом.
Подобная же проблема возникает при дифференциации двусторонних конечностей. Моя правая рука – асимметричный объект и формально зеркальное отражение моей левой руки. Но на свете встречаются редкие индивиды с чудовищным уродством – парой рук или разветвленной рукой с одной стороны тела. В таких случаях пара будет двусторонне симметричной системой. Одна из компонент будет правой, а другая левой, и обе будут расположены таким образом, что составляют зеркальное отражение друг друга.[43] Это обобщение было впервые опубликовано моим отцом в 1890-х годах и долго называлось «правилом Бейтсона». Он сумел доказать справедливость этого правила почти для всех подразделений животных, посредством исследований во всех музеях и многих частных коллекциях Европы и Америки. В частности, он собрал свыше ста случаев таких аберраций в ногах жуков.
Я заново рассмотрел эту историю и, опираясь на его исходные данные, пришел к выводу, что он ошибался в своей постановке вопроса. Он спрашивал: Что определяет эту добавочную симметрию? Между тем, надо было спрашивать: Что определяет потерю асимметрии?
|
|
|
Я предложил гипотезу, по которой уродливые формы производятся вследствие потери или забвения информации. Двусторонняя симметрия требует больше информации, чем радиальная симметрия, асимметрия же требует больше информации, чем двусторонняя симметрия. Асимметрия боковой конечности, такой, как рука, требует соответствующей ориентации в трехмерном пространстве. Направление к оборотной стороне руки должно быть отлично от направления к ладони; направление к большому пальцу должно быть отлично от направления к мизинцу, и направление к локтю должно быть отлично от направления к пальцам. Чтобы образовать правую руку – в отличие от левой – эти три направления должны быть надлежащим образом соединены. Если эти направления обращаются, как это происходит при отражении руки в зеркале, то получается обращенное изображение (см. Главу 3, раздел 9). Но если одна из дифференциаций потеряна или забыта, то конечность будет в состоянии достигнуть лишь двусторонней симметрии.
В этом случае постулат «ничто не происходит из ничего» принимает несколько более сложный вид: При потере одного различения из трех из асимметрии возникает двусторонняя симметрия.
гомология
Здесь я расстаюсь с проблемами индивидуальной генетики, соматического изменения и обучения, и непосредственными путями эволюции, чтобы рассмотреть результаты эволюции в более крупном масштабе. Я займусь теперь вопросом о том, что можно сказать о соответствующих процессах с точки зрения более широкой картины филогенеза.
Сравнительная анатомия имеет долгую историю. В течение по меньшей мере шестидесяти лет, от опубликования «Происхождения видов» до 1920-х годов, сравнительная анатомия была сосредоточена на взаимосвязях, не рассматривая процессов. Возможность построить филогенетические древа, как полагали, свидетельствовала в пользу теории Дарвина. Ископаемые данные были неизбежно весьма неполны, и в отсутствие прямых свидетельств о происхождении, анатомы проявляли ненасытный интерес к определенному классу сходства, именуемого гомологией. Гомология «доказывала» взаимосвязь, а взаимосвязь и была эволюцией.
Конечно, люди замечали формальное сходство живых существ по крайней мере с начала эволюции языка, соединявшего мою «руку» с вашей «рукой» и мою «голову» с «головой» рыбы. Но лишь гораздо позже возникло понимание необходимости объяснить такие формальные сходства. Даже в наше время большинство людей не удивляется сходству между двумя руками и не видит в этом никакой проблемы. Они не чувствуют и не усматривают никакой надобности в теории эволюции. Для мыслящих людей древности, и даже людей Возрождения формальное подобие живых существ было иллюстрацией связности Великой Цепи Бытия, и эти связи были для них логическими, а не генеалогическими связями.
|
|
|
Так или иначе, скачкообразное заключение от формального сходства к взаимосвязи скрывало ряд скачкообразных гипотез.
Признáем формальное сходство в тысячах случаев – между человеком и лошадью, омаром и крабом – и допустим, что в этих случаях формальное сходство – не только свидетельство, но попросту результат эволюционного отношения. Тогда мы можем рассмотреть вопрос, проливает ли природа сходства в этих случаях свет на процесс эволюции.
Спросим себя: что говорят нам гомологии о процессе эволюции? При сравнении нашего описания омара с нашим описанием краба мы обнаруживаем, что некоторые компоненты описаний остаются неизменными, а другие отличаются друг от друга. Поэтому первый шаг несомненно должен принять форму различения между разными видами изменения. Некоторые изменения будут подчеркнуты как более вероятные и более легкие; другие окажутся более трудными, и потому менее вероятными. В таких условиях медленно меняющиеся переменные будут отставать и могут стать ядром тех гомологий, на которых можно основывать более широкие гипотезы таксономии.
Но эта первая классификация изменений на быстрые и медленные и сама потребует объяснения.Чтó можно прибавить к нашему описанию эволюционного процесса, что могло бы, возможно, предсказать, какие переменные будут в самом деле медленно меняющимися и станут, тем самым, базисом гомологии?
Насколько мне известно, единственные начала такой классификации неявно содержатся в так называемой теории повторения.
Зародышем доктрины повторения было высказывание одного из пионеров эмбриологии, немца Карла Эрнста фон Бера в 1828 году о «законе соответствующих стадий». Он доказывал свой закон с помощью приема сравнения эмбрионов позвоночных разных непомеченных видов.
Я никак не в состоянии определить, к какому классу они принадлежат. Это могут быть ящерицы, или небольшие птицы, или очень молодые млекопитающие – столь полно сходство в способе формирования головы и туловища этих животных. Конечности еще отсутствуют, но если бы даже они существовали, на ранней стадии развития мы ничего бы не узнали, поскольку все возникает из одной и той же основной формы.[44]
Концепция «соответствующих стадий» фон Бера была позже разработана Эрнстом Геккелем, современником Дарвина, в теорию повторения и в часто оспаривавшееся утверждение, что «онтогенез повторяет филогенез». С тех пор были предложены весьма разнообразные формулировки этого положения. Самое осторожное из них – это, пожалуй, утверждение, что личинки или эмбрионы данного вида обычно более напоминают личинки или эмбрионы родственного вида, чем взрослые особи напоминают взрослые особи родственного вида. Но даже эта очень осторожная формулировка ставится под сомнение заметными исключениями.[45]
Но несмотря на эти исключения я склонен думать, что обобщение фон Бера составляет важный ключ к пониманию процесса эволюции. Это обобщение – верно оно или неверно – ставит важные вопросы о выживании не организмов, а признаков: Существует ли общий наибольший знаменатель тех переменных, которые становятся устойчивыми, и поэтому использовались зоологами в их поиске гомологии? Закон соответствующих стадий имеет то преимущество перед более поздними формулировками, что не претендует на филогенетические древеса, и даже краткая цитата, приведенная выше, содержит некоторые особенности, которых не заметит филогенетический изыскатель. Верно ли, что эмбриональные переменные более стойки, чем переменные взрослых?
Фон Бер сосредоточивается на высших позвоночных – ящерицах, птицах и млекопитающих – животных, эмбриология которых поддерживается и защищается либо наполненным пищей яйцом, либо материнской утробой. Но доказательства фон Бера просто не действовали бы, скажем, для личинок насекомых. Любой энтомолог, взглянув на не снабженную надписями выставку личинок жуков, может сразу сказать, к какому семейству принадлежит каждая личинка. Разнообразие личинок столь же удивительно, как и разнообразие взрослых особей.
Закон соответствующих стадий, по-видимому, верен не только для целых эмбрионов позвоночных, но также и для последовательности конечностей на самых ранних стадиях их развития. Так называемая серийная гомология разделяет с филогенетической гомологией то свойство, что, в общем, сходства предшествуют различиям. Вполне развитая клешня омара заметно отличается от ходячих конечностей на четырех других сегментах груди, но все конечности груди на их ранней стадии выглядят похоже.
Возможно, именно это и есть граница, до которой следует доводить обобщение фон Бера: можно утверждать, что, вообще говоря, сходство старше различия (и в филогенетическом, и в онтогенетическом смысле). Некоторым биологам это может показаться трюизмом, как если сказать, что в каждой ветвящейся системе две точки, близкие к точке ветвления, будут более сходны между собой, чем другие две точки, далекие от нее. Но этот кажущийся трюизм не будет справедлив для элементов периодической системы, и не обязательно будет справедлив в биологическом мире, произведенном намеренным творением. В действительности наш трюизм свидетельствует в пользу гипотезы, что организмы в самом деле связаны между собой, как точки или положения на ветвящемся дереве.
Обобщение, по которому сходство старше различия, – все еще весьма неполное объяснение наличия гомологии в тысячах примеров, во всем биологическом мире. Говоря, что сходства старше различий, мы только повторяем вопрос: «Почему некоторые признаки становятся основой гомологии?». Вопрос этот почти не меняется, принимая следующий вид: Почему некоторые признаки становятся старшими, сохраняясь дольше, и тем самым становятся основой гомологии?
Мы сталкиваемся здесь с проблемой выживания – не выживания видов или разновидностей, борющихся во враждебном мире других организмов, а с более тонкой проблемой выживания признаков (предметов описания), которые должны выжить и во внешнем окружении, и во внутреннем мире других признаков, во всей совокупности репродукции, эмбриологии и анатомии данного организма.
Почему же некоторые части описания, входящие в составленную ученым сложную систему описания всего организма, остаются в силе дольше других частей (в течение ряда поколений)? И есть ли совпадение, наложение или синонимия между частями и кусками описания и частями и кусками набора инструкций, определяющего онтогенез?
Если бы слон имел строение зубов и другие формальные признаки семейства Muridae*, он был бы мышью, несмотря на свой размер. В самом деле, даман, размером с кошку, близок к тому, чтобы быть гиппопотамом, а лев очень близок к тому, чтобы быть кошкой. Размер сам по себе, по-видимому, несуществен. Чтó важно – это форма. Но не так легко определить, что понимается под «формой» или «паттерном» в этом контексте.
Мы ищем критерии, позволяющие распознать признаки, которые, как можно думать, дольше остаются справедливыми в сумятице эволюционного процесса. Здесь выделяются два признака – два традиционных способа подразделения в обширном поле «различий». Существует дихотомия между паттерном и количеством и дихотомия между непрерывностью и разрывностью. Соединяются ли контрастирующие организмы непрерывным рядом шагов, или же отделены резким переходом? Затруднительно (хотя и возможно) представить себе постепенный переход между паттернами, а потому эти две дихотомии, по-видимому, перекрываются. Во всяком случае, можно ожидать, что теоретики, предпочитающие ссылаться на паттерны, будут также предпочитать теории, ссылающиеся на разрывность. (Но, конечно, подобные предпочтения, зависящие лишь от умонастроения отдельного ученого или от модного мнения, заслуживают порицания).
Я полагаю, что самые убедительные открытия в этом вопросе – это изящные доказательства зоолога Д’Арси Вентворта Томпсона, предложенные в начале двадцатого века. Как он показал, во многих случаях – пожалуй, во всех рассмотренных им случаях – две контрастирующих, но родственных формы животных имеют следующее общее свойство: если одна из этих форм изображена (скажем, в профиль) в декартовых ортогональных координатах (например, на разграфленной бумаге), то, при надлежащем наклоне или искажении, те же координаты подойдут и к другой форме. Все точки профиля второй формы совпадут с одноименными точками в искаженных координатах (см. рис. 9).
Примечательная особенность открытий Д’Арси Томпсона состоит в том, что в каждом случае искажение неожиданно просто, причем оно последовательным образом сохраняется в изображении всего животного. Наклон координат таков, что его можно описать некоторым простым математическим преобразованием.
Эта простота и последовательность должны несомненно означать, что различия между фенотипами, выявляемые методом Д’Арси Томпсона, представляют лишь немногие различия в генотипе (т.е. в небольшом числе генов).
Более того, последовательность искажения, распространяющаяся на все тело животного, по-видимому, указывает, что относящиеся к этому гены плейотропны (т.е. влияют на многие, может быть, на все части фенотипа), притом такими способами, которые в этом смысле гармонизированы по всему телу.
Дальнейшая интерпретация этих открытий не совсем проста, и сам Д’Арси Томпсон в этом не очень помогает. Его восхищает тот факт, что математика оказалась способной описать некоторые виды изменений.
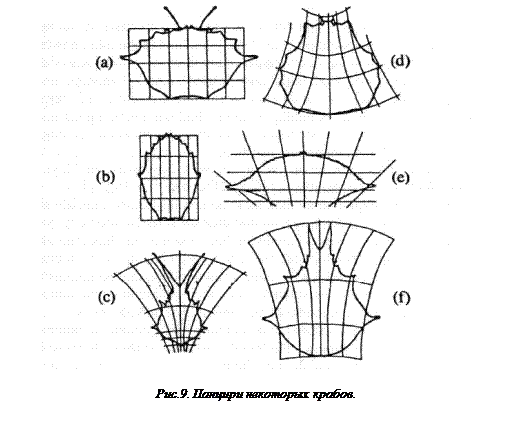
Рисунок заимствован из книги Д’Арси Томпсона «О росте и форме» (D’Arcy Tompson, On Growth and Form ), стр. 294. Cambridge University Press, 1961.
В этой связи любопытно отметить нынешний спор между сторонниками «синтетической» теории эволюции (т.е. нынешнего ортодоксального дарвинизма) и их противниками, «типологами». Например, Эрнст Майр высмеивает слепоту типологов: «Как показывает история, типолог нисколько не оценивает, и не способен оценить естественный отбор».[46] К сожалению, он не ссылается на источники своей типологии, применяемой им к его коллегам. Может быть, он слишком скромен, чтобы претендовать на эту честь? Или же, в этом случае, похожие узнают друг друга?
Но разве не все мы в душе типологи?
Во всяком случае, несомненно, есть много способов рассматривать формы животных. И поскольку мы принялись, в духе Платона, за исследование параллелизма между творческим мышлением и обширным разумным (mental) процессом, именуемым биологической эволюцией, то в каждом случае стóит спросить: Не представлен ли этот способ рассмотрения явлений, не параллелен ли он в некотором роде чему-то в системе организации самих явлений? Нет ли в каких-то генетических сообщениях и устойчивых признаках, определяющих фенотип, чего-то вроде синтаксиса (за неимением лучшего слова), отделяющего «типологическое» мышление от «синтетического»? Можем ли мы распознать среди множества сообщений, создающих и определяющих формы животных, некоторые более типологические и более синтетические?
Если поставить вопрос таким образом, то представляется, что Томпсон* глубоко прав, предлагая свою типологию. Старые рисунки Д’Арси Томпсона именно разделяют два вида коммуникаций внутри самого организма. Рисунки показывают, что у животных есть два вида признаков: (а) У них относительно устойчивые квази-топологические+ паттерны, которые понятным образом побудили ученых постулировать резкую разрывность в процессе эволюции. Эти признаки остаются неизменными под действием (б) относительно неустойчивых количественных свойств, видимых при сравнении смежных изображений.
Если начертить координатные линии, соответствующие квази-топологическим свойствам, то оказывается, что изменения менее устойчивых свойств представляются искажением координат.
В смысле рассматриваемого вопроса о гомологии, складывается впечатление, что в самом деле существуют разные виды свойств, и что филогенетическая гомология несомненно зависит от более устойчивых, квази-топологических паттернов.
7. приспособление и аддикция *
На языке эволюциониста «приспособление» означает приблизительно то же, что «план» на языке таких теологов, как Уильям Пейли,[47] «Доказательства» которого представляют собой объемистое собрание бросающихся в глаза примеров изящных специальных приспособлений животных к их образу жизни. Но я подозреваю, что и «адаптация», и «план» – понятия, вводящие в заблуждение.
Когда мы рассматриваем возникновение частных примеров приспособления – клешни краба, руки и глаза человека, и тому подобного – как центральную из всех проблем, подлежащих решению эволюциониста, мы искажаем и ограничиваем наш взгляд на эволюцию в целом. Как можно полагать, в результате нелепых сражений между ранними эволюционистами и церковью из всего обширного гераклитова потока эволюционного процесса были, по-видимому, выхвачены, в качестве предметов особого внимания, некоторые водовороты и заводи. Вследствие этого были в известной степени оставлены в стороне два великих стохастических процесса. Даже профессиональные биологи не видели, что – с более широкой точки зрения – эволюция столь же свободна от ценностей и столь же прекрасна, как танец Шивы, в котором все прекрасное и все уродливое, творение и разрушение выражены или сжаты в сложное симметричное движение.
Поместив термины приспособление и аддикция рядом друг с другом в заголовке этого раздела, я попытался исправить этот сентиментальный или, по крайней мере, чрезмерно оптимистический взгляд на эволюцию в целом. Вызывающие восхищение случаи приспособления, в которых природа кажется столь хитроумной и столь изобретательной, могут также быть начальными шагами к патологии и сверхспециализации. И все же трудно увидеть в клешне краба или в сетчатке человеческого глаза первые шаги к патологии.
По-видимому, мы должны спросить: Что же характеризует те приспособления, которые оказываются пагубными, и чем они отличаются от тех, которые кажутся благотворными и, подобно клешне краба, остаются благотворными в течение геологических эпох?
Это насущный вопрос, имеющий отношение к современным дилеммам нашей цивилизации. Во времена Дарвина благотворным казалось каждое изобретение, но теперь дело обстоит иначе. В двадцатом столетии изощренные глаза рассматривают каждое изобретение с подозрительностью, сомневаясь в том, всегда ли слепые стохастические процессы все вместе действуют во благо.
Мы остро нуждаемся в науке, способной анализировать всю совокупоность приспособления-аддикции на всех уровнях. Может быть, экология начинает становиться такой наукой, хотя экологи далеко не в состоянии сказать нам, как выбраться из гонки атомных вооружений.
В принципе, ни случайные генетические изменения, сопровождаемые естественным отбором, ни случайные процессы мышления по методу проб и ошибок, сопровождаемые избирательным подкреплением, не обязательно идут на благо виду или индивиду. А на социальном уровне еще не ясно, обязательно ли изобретения и стратегии, вознаграждающиеся для индивида, имеют ценность для выживания общества; и обратно, обязательно ли образ действий, который предпочли бы влиятельные представители общества, имеет ценность для выживания индивида.
Можно привести большое число примеров, подсказывающих, что вера в естественный отбор или в laissez-faire* очевидным образом наивна:
а. Остаток системы изменяется и закрепляет новшество, чтобы сделать его необратимым.
б. Взаимодействие с другими видами или индивидами приводит к изменению контекста, при котором становится необходимым дальнейшее новшество того же рода, и таким образом в системе начинается эскалация, или она идет вразнос.
в. Новшество вызывает другие изменения в системе, вынуждающие отказ от других приспособлений.
г. Уменьшается гибкость системы (т.е. ее положительная энтропия).
д. Приспособленный вид оказывается в столь благоприятных условиях, что разрушает свою экологическую нишу какой-нибудь формой чрезмерного выпаса.
е. Нечто, казавшееся желательным в ближайшей перспективе, на длительный период оказывается пагубным.
ж. После новшества вид или индивид начинает вести себя так, как будто он больше не находится в частичной зависимости от смежных видов и индивидов.
з. Посредством аддикции новшество втягивается в стремление сохранить некую постоянную скорость изменения. Социальная аддикция гонки вооружений не отличается в своей сущности от индивидуальной аддикции к наркотикам. Здравый смысл всегда побуждает наркомана снова уколоться. И так далее.
В итоге, в каждом из этих бедствий можно обнаружить ошибку в определении логического типа. Вопреки немедленному выигрышу на некотором логическом уровне, знак обращается и выгода превращается в бедствие в некотором другом, большем и более длительном контексте.
У нас нет никакого систематического знания о динамике этих процессов.
|
|
|
