Регуляция клеточного цикла у млекопитающих.
На предыдущей схеме рассмотрены только конечные звенья цепи событий, заканчивающихся делением клетки. Однако, как уже говорилось, деление клетки обязательно связано с репликацией ДНК. Следовательно, должны существовать механизмы, регулирующие запуск синтеза ДНК, а этому должны предшествовать события G1-периода, подготавливающие начало S-периода. Оказалось, что комплексы Cdk—митотический циклин В — это только конечный этап регуляции клеточного цикла. На самом же деле от начала вхождения в клеточный цикл до его завершения в клетке работает каскад комплексов Cdk - циклин. Так, у дрожжей один и тот же Cdk отвечает за прохождение цикла, но на разных его стадиях он взаимодействует с разными циклинами, характерными для каждой стадии клеточного цикла.
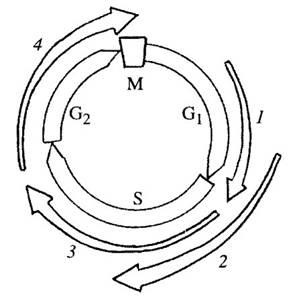
У млекопитающих в реализации всего цикла участвуют девять различных циклинов и семь разных Cdk (рис. 351). Но что является пусковым механизмом для вхождения клеток в цикл из состояния покоя, из G0-стадии? Мы видели, что для активации клеточного цикла ооцита X. laevis необходимо первоначальное воздействие гормона прогестерона, который в данном случае является фактором роста (ФР, или GF) или пролиферации.
|
| Рис. 351. Участие в клеточном цикле млекопитающих различных циклинов и циклинзависимых киназ
1 — циклин Д + Cdk 4, Cdk 6; 2 — циклин Е + Cdk 2; 3 — циклин А + Cdk 2; 4 — циклин В/А + Cdk 1
|
Было найдено, что существует множество факторов роста, побуждающих клетки к размножению. Они могут быть собственными продуктами данных клеток (аутокринная стимуляция) или других соседних (паракринная стимуляция), или даже клеток других органов (гормональная стимуляция). Так, фактор роста из тромбоцитов (PDGF) стимулирует пролиферацию клеток соединительной ткани, эпидермальный ФР (EGF) — размножение многих типов клеток, работает как сигнальный белок при эмбриональном развитии; ФР нервов (NGF) вызывает рост отдельных типов нейронов; эритропоэтин — пролиферацию предшественников эритроцитов и т.д.
Хорошим примером зависимости пролиферации клеток от факторов роста могут служить клеточные культуры. Было найдено, что для размножения многих клеточных культур, кроме питательных сред, необходимо добавление сывороток крови или эмбриональных экстрактов. Оказалось, что именно в этих добавках содержатся ФР. Без них клетки переходят в G0-стадию, а затем погибают. Для роста нервных клеток в культуре требуется добавление в среду фактора роста нервов (NGF).
Эти разные ФР связываются на поверхности клеток со своими рецепторами и передают сигнал через вторичные мессенджеры (например, цАМФ) на систему внутриклеточного каскада протеинкиназ (фосфорилаз), связанных с запуском клеточного цикла. Сначала активируются гены раннего ответа, белки которых индуцируют транскрипцию генов отложенного ответа, некоторые из них сами являются факторами транскрипции, а также индуцируют синтез ряда циклинов и Cdk, которые отсутствовали в G0-периоде.
|
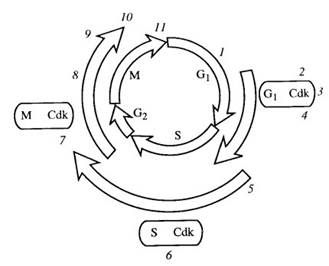
| Рис. 352. Общая схема регуляции цикла эукариотической клетки
1 — сборка ДНК-репликативных коммлексов; 2 — G1-Cdk-комплекс инактивирует APC (комплекс, стимулирующий анафазу); 3 — G1-Cdk-комплекс активирует транскрипцию Cdk-комплекса S-фазы; 4 — G1-Cdk-комплекс фосфорилирует ингибитор Cdk-комплекса S-фазы; 5 — деградация этого ингибитора; 6 — Cdk-комплекс S-фазы активирует репликативные комплексы; 7 — митотический Cdk-комплекс (М-Cdk) активирует конденсацию хромосом, разрыв ядерной оболочки, сборку веретена деления; 8 — активирование АРС; 9 — АРС разрушает анафазный ингибитор, что приводит к расхождению хромосом в анафазе; 10 — АРС вызывает деградацию митотического циклина с помощью протеосом; 11 — телофаза и цитокинез
|
Так, вначале синтезируются белки Cdk и циклины, характерные для G1-стадии, затем для S-фазы и потом для митоза (рис. 352). В G1-стадии комплекс Cdk—циклин (G1—CDK) фосфорилирует факторы транскрипции, необходимые для активации экспрессии генов, ответственных за образование синтетического комплекса S-CDK, который после образования инактивируется специальным ингибитором. В конце G1-периода комплекс G1-CDK фосфорилирует этот ингибитор, который отделяется от комплекса S—CDK, тем самым его активируя. При этом в первоначальном комплексе G1—CDK циклин деградирует. Активированный S-CDK-комплекс индуцирует S-фазу, фосфорилируя белки регуляторного участка ДНК, связанных с точками начала репликации. Затем циклин в этом комплексе также деградирует. После активации S-периода происходит репликация ДНК. Во время S-периода и в начале G2-периода происходит синтез нового митотического комплекса M-CDK, определяющего вхождение клетки в митоз. Однако до окончания синтеза ДНК он находится в неактивном состоянии и активируется путем дефосфорилирования. После активации этого комплекса он участвует в фосфорилировании белков хроматина, что приводит к конденсации хромосом, белков ламины, которые деполимеризуются, и при этом разрушается ядерная оболочка, фосфорилирует ряд белков, ассоциированных с микротрубочками при образовании веретена деления. После ассоциации микротрубочек с хромосомами происходит активация АРС (комплекс стимуляции анафазы), деградация когезинов, вслед за чем наступает анафаза и активация протеолиза митотических циклинов. После расхождения хромосом и цитотомии в раннем G1-периоде следующего цикла новые комплексы G1-CDK фосфорилируют АРС, инактивируя их, что способствует впоследствии накоплению митотических циклинов.
Воспользуйтесь поиском по сайту:



