 |


Генетические аномалии строения коры у мышей линии reeler
|
|
|
|
Другой пример того, как определяется дальнейшее развитие нейрона до того, как он достигнет своего окончательного расположения, обнаружен у мутантных мышей линии reeler (названных так из-за своей неровной походки). В развивающейся коре мышей reeler более молодые нейроны не мигрируют сквозь слои других клеток. Таким образом, их расположение друг относительно друга у взрослых противоположное: нейроны, образованные раньше, располагаются в более поверхностных слоях, а нейроны, рожденные позже, — в более глубоких. Несмотря на аномальное расположение, нейроны имеют правильную исходную морфологию (определяемую моментом рождения) и образуют между собой связи в зависимости от времени образования. Получается, что морфология корковых нейронов и природа их синаптических взаимодействий определяются в момент рождения нейрона и могут проявляться независимо от его расположения. Такой reeler фенотип является результатом большого количества мелких изменений в результате мутации в коре и других областях мозга.
Продуктом экспрессии гена reeler является большой гликопротеин внеклеточного матрикса, называемый reelin. Он не экспрессируется в кортикальных клетках, не мигрирующих правильным образом, а в основном экспрессируется в клетках Кахаля—Ретциуса в краевой зоне коры. Нарушения экспрессии других генов в кортикальных клетках могут приводить к сходным с reeler фенотипам проявления. Например, мутация цитоплазматического белка, содержащего фосфотирозин (cytoplasmic phosphotyrosine-containing protein), названного Disabled-1, также приводит к reeler фенотипу, как и генетическое выключение (knock-out) рецептора липопротеинов низкой плотности вместе с рецепторами 2 типа к аполипопротеину Е. На основании этого можно судить о том, что эти три белка являются частью внутриклеточного механизма, который позволяет кортикальным клеткам распознавать сигнал с reelin-белка, передающийся через внеклеточный матрикс, и использовать этот сигнал для установления правильной структуры слоев коры. Большое количество других генетических мальформаций в коре ГМ человека связано с нарушением миграции нейронов.
|
|
|
Влияние локальных сигналов на корковую архитектуру
Примером способности корковых нейронов изменять свое строение после того, как они мигрировали прочь от вентрикулярной зоны, является эксперимент, в котором кусочек развивающейся зрительной коры был пересажен в область представления вибрисс соматосенсорной коры крысы, . Нейроны в трансплантанте реорганизовались и сформировали четко различимую колончатую структуру, характерную для их нового месторасположения, образуя фенотип, не характерный для зрительной коры. Таким образом, внутренние влияния могут определять особенности фенотипов корковых нейронов.
Гормональный контроль за развитием нервной системы
В некоторых областях ЦНС развитие нейронов находится под гормональным контролем. Это особенно заметно в областях мозга, определяющих сексуальное поведение. Например, у таких певчих птиц, как канарейки, ядро высшего вокального центра (ВВЦ, high vocal center, HVC) играет ключевую роль в приобретении и запоминании песни — что является исключительно мужским поведением. Эта зона мозга больше развита у самцов, чем у самок (рис. 3.). Однако пение можно вызывать даже у взрослых самок введением тестостерона, в результате чего ядро ВВЦ и другие структуры, связанные с воспроизведением песни, увеличиваются.
Ядра ВВЦ у взрослых самцов и самок канарейки являются особенными центрами, так как там происходит постоянный кругооборот нейронов). Образование (recruitment) новых нейронов ВВЦ у самцов происходит максимально осенью и весной, сразу после пика гибели старых нейронов. Таким образом, в это время происходит модификация их песен для нового брачного сезона. Период гибели нейронов совпадает с падением уровня тестостерона, а образование новых — с его пиком. Назначение тестостерона самкам приводит к увеличению образования новых нейронов в ВВЦ, которые, как их предшественники, получают определенные синаптические сигналы и посылают свои аксоны к определенным мишеням. Эти удивительные наблюдениям показывают, что в зрелом мозге возможно не только образование новых нейронов, но также их включение в сложно устроенную и функционирующую систему, что приводит к перемоделированию такого свойственного птицам поведения, как их песни.
|
|
|
Стволовые нервные клетки
ВВЦ певчих птиц является не единственным примером того, когда новые нейроны включаются в состав нервной системы взрослых животных.
| Рис. 3.. Половой диморфизм в головном мозге птиц. Схематическая диаграмма основных зон мозга и путей, участвующих в формировании песни у певчих птиц. Высший вокальный центр (HVC), крепкое ядро архистриатума (RA) и подъязычное ядро формируют задний вокальный моторный путь (показано черным). HVC, зона X, медикальное дорзолатеральное ядро тала муса (DLM) и латеральное магноцеллюлярное ядро переднего неостриатума (LMAN) образуют передний путь (показано серым). HVC, подъязычное ядро и RA имеют значительно большие размеры у самцов; зона X не обнаруживается в мозге самок зябликов. Fig. 23.14. Sexual Dimorphism in the Avian Brain. Schematic diagram of the major brain areas and pathways involved in production of song in songbirds. | 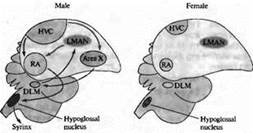
|
Нейроны также постоянно обновляются в гиппокампе и обонятельной луковице у взрослых млекопитающих. Однако как образуются эти клетки? Нервные стволовые клетки, которые обладают способностью к самообновлению, были выделены из стенок желудочков и из гиппокампа взрослого мозга и было прослежено их размножение in vitro. Эти клетки могут дифференцироваться в нейроны, олигодендроциты и астроциты. Нейроны, постоянно добавляющиеся в обонятельной луковице in vivo, происходят из медленно делящихся стволовых клеток самого внутреннего, эпендимального слоя стенок боковых желудочков (остаток начальной вентрикулярной зоны). Один из потомков каждого клеточного деления направляется в субэпендимальную зону, чтобы стать там клеткой-предшественницей. Клетка-предшественница быстро делится, образуя незрелые нейроны, которые мигрируют в ростральном направлении в область луковицы, где они дифференцируются в интернейроны и интегрируются в имеющиеся сети. При повреждении ЦНС клетка-потомок стволовых клеток становится астроцитом, а не нейроном, мигрирует в область повреждения и принимает участие в образовании рубца в области повреждения. Таким образом, судьба потомка стволовой клетки у взрослых может быть изменена под влиянием внешних сигналов.
|
|
|
Стволовые нервные клетки, таким образом, представляют собой популяцию клеток, способных размножаться в культуре и подвергаться дифференцировке либо в глиальные клетки, либо в нейроны. Это обеспечивает возможность использования этих клеток в качестве резерва нейронов или глиальных клеток при лечении заболеваний нервной системы, при которых наблюдается гибель клеток ГМ или их дисфункция.
|
|
|
