 |


Управляемые каналы клеточной мембраны
|
|
|
|
 Только что описанные каналы, через которые проходят ионы калия, открыты всегда: и в состоянии покоя, и при возбуждении клетки – они мало зависят от внешних условий и поэтому являются каналами пассивного типа. В противоположность этому существуют управляемые ионные каналы, большинство из которых в состоянии покоя клетки закрыты, а для того, чтобы их открыть, необходимо каким-то образом на них подействовать. Следовательно, такие каналы являются управляемыми, а в зависимости от способа управления их подразделяют на три типа: потенциалзависимые; хемозависимые; управляемые механически.
Только что описанные каналы, через которые проходят ионы калия, открыты всегда: и в состоянии покоя, и при возбуждении клетки – они мало зависят от внешних условий и поэтому являются каналами пассивного типа. В противоположность этому существуют управляемые ионные каналы, большинство из которых в состоянии покоя клетки закрыты, а для того, чтобы их открыть, необходимо каким-то образом на них подействовать. Следовательно, такие каналы являются управляемыми, а в зависимости от способа управления их подразделяют на три типа: потенциалзависимые; хемозависимые; управляемые механически.
Потенциалзависимые каналы открываются или закрываются в связи с определёнными изменениями мембранного потенциала. Например, натриевые каналы в состоянии покоя закрыты, но, если мембранный потенциал уменьшится до критического значения, они открываются. Если деполяризация продолжится до положительного значения мембранного потенциала (т.е. на внутренней части мембраны окажется больше положительных зарядов, чем на наружной), то каналы закроются.
Хемозависимые каналы открываются вследствие присоединения нейромедиатора к выступающей наружу гликопротеиновой рецепторной области канального белка – такой тип каналов используется в синапсах (рис. 4.4). Механически управляемые каналы характерны для чувствительных окончаний нейронов, реагирующих на растяжение и давление. Эти каналы особым способом связаны с цитоскелетом, что и приводит их к открытию при деформации клетки.
Сам момент открытия канала – всего лишь мгновение, длящееся миллионные доли секунды. Но и в открытом состоянии каналы находятся недолго – лишь несколько миллисекунд, после чего стремительно закрываются. Однако пропускная способность открытого канала поразительна: ток ионов происходит со скоростью до 100 000 000 ионов/с, что можно сравнить только с деятельностью самых "быстрых" ферментов, как, например, карбоангидраза, катализирующая образование и дегидратацию углекислоты в эритроцитах крови.
|
|
|
Кроме открытого и закрытого конформационных состояний каналы могут стать инактивированными: это значит, что они закрыты, но не подчиняются, как обычно, действию механизмов управления и не открываются. Состояние инактивации наблюдается немедленно после закрытия каналов, продолжается несколько мс и контролируется особыми субъединицами или особыми регионами белковой молекулы. Во время инактивации каналов клетка перестаёт отвечать на возбуждающие её стимулы, что определяется термином рефрактерность, т.е. временная невозбудимость.
Ионные каналы имеются в мембране любой клетки организма, но в мышечных и особенно в нервных клетках их плотность намного больше, чем в клетках других тканей. В нейронах, кроме высокой плотности каналов, обнаружено и их большое разнообразие. Это не случайно, поскольку именно каналы определяют условия возникновения электрических сигналов, характер самих сигналов, скорость их проведения и т.п., что собственно и позволяет нейронам выполнять свою главную задачу: принимать, перерабатывать и передавать информацию.
Потенциал действия
Механизм потенциала действия достаточно сложен, и его удобнее рассматривать по фазам. В ПД различают фазу деполяризации, фазу реполяризации и следовые потенциалы.
 Механизм фазы деполяризации: действие раздражителя приводит к неспецифическому ответу клетки в виде открывания натриевых каналов, что приводит к деполяризации мембраны. Это в свою очередь облегчает открывание все большего количества натриевых каналов, что еще сильнее деполяризует мембрану. Таким образом, деполяризация мембраны достигает определенной степени, при которой открываются все натриевые каналы (рис. 7А). Эта степень деполяризации называется критическим уровнем деполяризации (КУД). При этом натрий начинает быстро проникать в клетку, доводя разность потенциалов между внутренней и наружной поверхностью мембраны до 0, а затем наступает перезарядка мембраны (инверсия потенциала), то есть внутренняя ее поверхность становится положительна заряженной относительно наружной. Но поступление ионов натрия в клетку не бесконечно. Оно ограничивается натриевой инактивацией (каналы не могут быть открытыми длительное время). Кроме того, проникшие в клетку ионы натрия создают электрическое поле, препятствующее дальнейшему поступлению натрия (рис.7Б).
Механизм фазы деполяризации: действие раздражителя приводит к неспецифическому ответу клетки в виде открывания натриевых каналов, что приводит к деполяризации мембраны. Это в свою очередь облегчает открывание все большего количества натриевых каналов, что еще сильнее деполяризует мембрану. Таким образом, деполяризация мембраны достигает определенной степени, при которой открываются все натриевые каналы (рис. 7А). Эта степень деполяризации называется критическим уровнем деполяризации (КУД). При этом натрий начинает быстро проникать в клетку, доводя разность потенциалов между внутренней и наружной поверхностью мембраны до 0, а затем наступает перезарядка мембраны (инверсия потенциала), то есть внутренняя ее поверхность становится положительна заряженной относительно наружной. Но поступление ионов натрия в клетку не бесконечно. Оно ограничивается натриевой инактивацией (каналы не могут быть открытыми длительное время). Кроме того, проникшие в клетку ионы натрия создают электрическое поле, препятствующее дальнейшему поступлению натрия (рис.7Б).
|
|
|
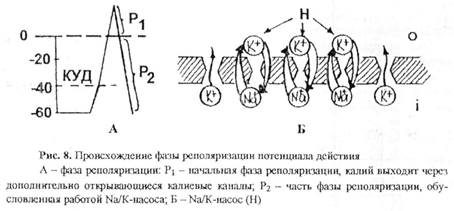
 Механизм фазы реполяризации: в ответ на поступление ионов натрия в клетку быстро включаются два механизма, возвращающие исходную степень поляризации мембраны (рис 8). Во-первых, открываются те калиевые каналы, которые были закрыты в состоянии покоя, и калий выходит из клетки в значительно большем объеме, что уменьшает степень деполяризации поверхностной мембраны клетки. Во-вторых, активируется работа натрий-калиевого насоса, возвращающего исходную ионную асимметрию по обе стороны поверхностной мембраны клетки. Таким образом, происходит восстановление МПП.
Механизм фазы реполяризации: в ответ на поступление ионов натрия в клетку быстро включаются два механизма, возвращающие исходную степень поляризации мембраны (рис 8). Во-первых, открываются те калиевые каналы, которые были закрыты в состоянии покоя, и калий выходит из клетки в значительно большем объеме, что уменьшает степень деполяризации поверхностной мембраны клетки. Во-вторых, активируется работа натрий-калиевого насоса, возвращающего исходную ионную асимметрию по обе стороны поверхностной мембраны клетки. Таким образом, происходит восстановление МПП.
В идеале следовых потенциалов не должно быть, так как фаза реполяризации возвращает клетку в состояние покоя с исходным МПП и исходной возбудимостью. Но в реальности фаза реполяризации может растянуться по времени из-за недостаточно активной работы Na/K-насоса и возникает следовая деполяризация (отрицательный следовой потенциал) (рис. 9А). Напротив, если работа Na/K-насоса усилена, то возникает следовая гиперполяризации (положительный следовой потенциал) (рис. 9Б). Иногда эти потенциалы следуют друг за другом (рис. 9В).
На допороговые раздражители, не превышающие но своей силе 50% от величины порогового раздражителя, клетка не реагирует вообще. Эти раздражители слишком слабы для того, чтобы в ответ на них на поверхностной мембране клетки дополнительно открывались натриевые каналы.
 На допороговые раздражители, составляющие по своей силе 50% и более от величины порогового раздражителя, в мембране клетки дополнительно открываются закрытые в состоянии покоя натриевые каналы. При этом возникает деполяризация поверхностной мембраны клетки, и она будет тем больше, чем сильнее действующий допороговый раздражитель. Эту деполяризацию обозначают как «локальный ответ».
На допороговые раздражители, составляющие по своей силе 50% и более от величины порогового раздражителя, в мембране клетки дополнительно открываются закрытые в состоянии покоя натриевые каналы. При этом возникает деполяризация поверхностной мембраны клетки, и она будет тем больше, чем сильнее действующий допороговый раздражитель. Эту деполяризацию обозначают как «локальный ответ».
|
|
|
 Пороговый потенциал – это часть мембранного потенциала покоя (рис. 11), на величину которой надо деполяризовать поверхностную мембрану клетки, чтобы достичь КУД (то есть, чтобы возникло возбуждение).
Пороговый потенциал – это часть мембранного потенциала покоя (рис. 11), на величину которой надо деполяризовать поверхностную мембрану клетки, чтобы достичь КУД (то есть, чтобы возникло возбуждение).
При действии допороговых раздражителей, составляющих менее 50% от величины порога раздражения, возбудимость клетки не изменяется (рис. 12, раздражители 1 и 2), так как не изменяется пороговый потенциал (исключение составляет постоянный ток).
При действии допороговых раздражителей, составляющих 50% и более от величины порога раздражения (рис.12, раздражители 3, 4, 5). Возбудимость клетки всегда повышается, т.к. пороговый потенциал уменьшается. Причем возбудимость будет тем больше, чем больше сила допорогового раздражителя.
 Изменения возбудимости будут носить фазный характер в соответствии с фазами потенциала действия, который возникнет в том и другом случае (рис. 13) Сразу после действия раздражителя (пока деполяризация не достигла критического уровня) возбудимость будет возрастать, т.к. пороговый потенциал будет уменьшаться вплоть до достижения КУД (рис. 13А, а). При достижении КУД возбудимость клетки исчезнет, т.к. все натриевые каналы будут открыты, и клетке нечем будет отвечать на действие даже очень сильного раздражителя (рис. 13А, б). Эта фаза называется абсолютной рефрактерностъю, то есть ткань в это время полностью невозбудима. Она будет сопровождать всю фазу деполяризации и начальный период фазы реполяризации, который обусловлен усиленным выходом калия из клетки.
Изменения возбудимости будут носить фазный характер в соответствии с фазами потенциала действия, который возникнет в том и другом случае (рис. 13) Сразу после действия раздражителя (пока деполяризация не достигла критического уровня) возбудимость будет возрастать, т.к. пороговый потенциал будет уменьшаться вплоть до достижения КУД (рис. 13А, а). При достижении КУД возбудимость клетки исчезнет, т.к. все натриевые каналы будут открыты, и клетке нечем будет отвечать на действие даже очень сильного раздражителя (рис. 13А, б). Эта фаза называется абсолютной рефрактерностъю, то есть ткань в это время полностью невозбудима. Она будет сопровождать всю фазу деполяризации и начальный период фазы реполяризации, который обусловлен усиленным выходом калия из клетки.
После активизации Na/K-насоса возбудимость клетки начинает восстанавливаться до исходного уровня. Эта фаза носит название относительной рефрактерности, то есть пониженной возбудимости (рис. 13А, в). Она сопровождает фазу реполяризации до ее окончания. В этот промежуток времени достаточно сильный раздражитель (сверхпороговый) может вызвать повторный потенциал действия.
|
|
|
В фазу отрицательного следового потенциала возбудимость будет повышена, так как пороговый потенциал в это время снижен (рис. 13Б, г). Напротив, в фазу положительного следового потенциала возбудимость будет снижена, так как пороговый потенциал в это время становится больше, чем в состоянии покоя (рис. 13В, г).
Благодаря фазе абсолютной рефрактерности один ПД отделяется от другого, не сливаясь с предыдущим. Это обеспечивает возможность кодирования информации, которое осуществляется нервной клеткой для реализации регулирующих влияний на другие возбудимые клетки. Кроме того, благодаря фазе абсолютной рефрактерности, осуществляется одностороннее проведение возбуждения.
Во всех тканях, обладающих проводимостью, она отличается по скорости проведения возбуждения, которая зависит от плотности расположения натриевых каналов на поверхностной мембране клетки. Чем она больше, тем выше скорость проведения возбуждения. В нервных волокнах на скорость проведения возбуждения существенно влияют его толщина и степень миелинизации. В связи с этим различают волокна типа А, В и С. Например, в волокнах типа А (диаметр 12-22 мкм, полностью покрыты миелиновой оболочной) скорость проведения наибольшая – 80-120 м/сек. Эти волокна проводят возбуждение от альфа-мотонейронов спинного мозга до миоцитов скелетных мышц. В волокнах типа С (диаметр – около 1 мкм, миелиновой оболочки не имеют) скорость проведения возбуждения наименьшая – 0,5-3 м/сек. Такие волокна проводят возбуждение, например, в постганглионарных волокнах вегетативной нервной системы.
|
|
|
