 |


Теория равновесия МакАртура и Уилсона.
|
|
|
|
Для объяснения этих странных закономерностей в 1967 г. МакАртур и Уилсон предложили "теорию равновесия". Суть этой теории:
количество видов острова определяется равновесием между иммиграцией и вымиранием, причем равновесие динамично - виды постоянно вымирают и замещаются иммигрантами из соседних экосистем (теми же или другими).
 Согласно этой теории, причиной увеличения количества видов является иммиграция. Для перемещающихся случайным образом организмов (с ветром, течением и т.д.) крупные и близкие острова являются более удачными "мишенями", чем мелкие и далекие. По мере заселения острова доля, которую вносит новый вид-иммигрант в изменение богатства видового состава, последовательно сокращается. Это видно из таблицы и графика (рис.3.5):
Согласно этой теории, причиной увеличения количества видов является иммиграция. Для перемещающихся случайным образом организмов (с ветром, течением и т.д.) крупные и близкие острова являются более удачными "мишенями", чем мелкие и далекие. По мере заселения острова доля, которую вносит новый вид-иммигрант в изменение богатства видового состава, последовательно сокращается. Это видно из таблицы и графика (рис.3.5):
| появление новых видов в сообществе | изменение видового состава |
| 1-ый иммигрант | на 100% |
| 2-ой иммигрант | на 50% |
| 3-ий | на 33% |
| 4-ый | на 25% |
| 5-ый | на 20% |
По оси ОУ - скорость иммиграции (число видов /год).
1 - крупный или близкий остров, 2 - мелкий или далекий остров.
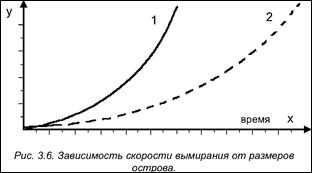
В результате постоянного вымирания количество видов, населяющих остров, не может расти до бесконечности. Ресурсы островной экосистемы во многом зависят от ее размеров. Этим объясняется высокая скорость вымирания организмов на мелких островах и низкая – на крупных. Темп вымирания растет по мере увеличения количества видов. Удаленность острова не влияет на скорость вымирания видов (рис. 3.6).
По оси ОУ - скорость вымирания (число видов /год).
1 - мелкий остров, 2 - крупный остров.
 В тот момент, когда количество новых видов-иммигрантов будет равно количеству вымирающих видов, установится динамическое равновесие, и число видов на острове станет относительно постоянным. Момент, когда будет достигнуто это равновесие, можно увидеть на графике. Для этого необходимо на одних и тех же координатных осях и в одном масштабе вычертить графики скорости иммиграции и вымирания, характерные для данного острова согласно теории Мак-Артура - Уилсона. Точка их пересечения характеризует число видов и время, при которых будет достигнуто динамическое равновесие богатства видового состава.
В тот момент, когда количество новых видов-иммигрантов будет равно количеству вымирающих видов, установится динамическое равновесие, и число видов на острове станет относительно постоянным. Момент, когда будет достигнуто это равновесие, можно увидеть на графике. Для этого необходимо на одних и тех же координатных осях и в одном масштабе вычертить графики скорости иммиграции и вымирания, характерные для данного острова согласно теории Мак-Артура - Уилсона. Точка их пересечения характеризует число видов и время, при которых будет достигнуто динамическое равновесие богатства видового состава.
|
|
|
График, иллюстрирующий равновесную теорию
островного эффекта (Мак-Артура и Уилсона).
Следует заметить, что равновесная теория значительно упрощает реальные процессы развития изолированных экосистем. Она не учитывает эволюционных процессов в рамках самой экосистемы, численные соотношения различных функциональных групп и разнообразие условий существования на островах равной площади и удаленности, но разных типов (вулканические, коралловые, материковые). Но это очень удобная схема, позволяющая объяснить островной эффект и рассчитать приблизительное видовое богатство любого острова. Для более детального анализа потребуется учет множества дополнительных параметров.
Дисгармонию видового состава и незаполненность экологических ниш изолятов можно объяснить несколькими причинами:
Во-первых, разные виды обладают принципиально отличающимися способностями к расселению. Многие птицы, летающие насекомые, споры грибов и бактерий, переносимые ветром, легко пересекают непригодные для жизни территории, которые являются непреодолимыми барьерами для менее подвижных видов. Эти препятствия снижают возможности иммиграции многих организмов.
Во-вторых, взаимная зависимость организмов друг от друга снижает вероятность поселения на новой территории консументов второго и выше порядков (кроме паразитов). Это легко понять: хищники на острове без жертв выжить не могут, а жертвы без хищников размножаются очень интенсивно. В таком случае численность жертв регулируется внутривидовой конкуренцией и паразитарными организмами, что может приводить к резким колебаниям численности и даже к вымиранию жертв.
|
|
|
Наконец, разная предрасположенность видов к вымиранию накладывает свой отпечаток на видовой состав сообщества. Особенно уязвимы виды крупных организмов, для которых характерна низкая плотность популяций. Последними двумя причинами объясняется тот факт, что на островах обычно нет крупных хищников. Дисгармония видового состава определяет неустойчивость экосистем изолятов и постоянное вымирание части видов, предсказанное равновесной теорией.
Незначительное число реликтовых видов, для которых условия изолята оказались благоприятны, может сохраниться. Им не приходится выдерживать серьезной конкуренции с эволюционно более молодыми претендентами на ту же экологическую нишу, а малочисленные "новоселы-иммигранты" сами оказываются в условиях весьма жесткой конкуренции. Но, если иммигранты выдержат первоначальную конкуренцию, реликтовые формы обречены. Этим опасна интродукция для изолированных сообществ. На островах, как и в любых экосистемах, протекают эволюционные процессы. Изоляция способствует накоплению и закреплению новых признаков в популяциях "островитян" и видообразованию. Появившиеся на островах эволюционным путем виды строго эндемичны по причине изолированности экосистем. Нарушение изоляции вследствие иммиграции родственных организмов из других экосистем замедляет эволюционный процесс. Уильямсон предложил выделять среди островов два экологических типа:
- океанические - эволюция видов идет быстрее иммиграции (высокая степень изоляции),
- континентальные - эволюция видов идет медленнее иммиграции (низкая степень изоляции).
Сочетание реликтовых видов и эволюционно молодых эндемичных форм – характерная черта океанических островов. Но и эта классификация довольно условна: разные виды имеют разные миграционные способности, и один и тот же остров можно назвать океаническим или континентальным в зависимости от анализируемой группы организмов.
|
|
|
Краткое содержание § 17
Экосистемы островов и других изолятов обладают рядом особенностей, не свойственных сообществам других экосистем: бедность видового состава, дисгармония видового состава (например, многие изоляты лишены крупных хищников), незаполненность экологических ниш, сочетание древних и эволюционно молодых эндемичных видов. При этом наблюдаются следующие закономерности:
- если два острова отличаются по площади в 10 раз, то число видов, населяющих меньший остров, в среднем вдвое меньше (при прочих сходных условиях);
- при возрастании удаленности острова на 2600 км количество видов в среднем сокращается вдвое.
Для объяснения этих закономерностей предложена "теория равновесия": количество видов острова определяется равновесием между иммиграцией и вымиранием, причем равновесие динамично: виды постоянно вымирают и замещаются иммигрантами. Таким образом, равновесная теория объясняет особенности экосистем изолятов их неустойчивостью и постоянным вымиранием части видов.
Задания.
1. Можно ли ожидать, что небольшой заповедник сохранит богатство видового состава первичной экосистемы, если прилегающие территории занимаются только под жилые массивы и экологически чистые производства? Зачем при организации мелких заповедников отдельные территории соединяют "заповедными коридорами"?
2. Постройте в одних координатных осях условные графики, отражающие:
- изменение скорости иммиграции видов на крупный удаленный остров,
- изменение скорости иммиграции на близкий остров того же размера,
- изменение скорости вымирания видов на острове такого же размера.
Сравните время достижения и относительное число видов точки динамического равновесия.
3. Выполните построения, аналогичные предыдущим, но для маленьких островов. Сравните результаты, полученные в этих двух заданиях. Сделайте вывод о причинах влияния размеров и удаленности островов на видовое богатство.
 4. Даны два острова одного возраста, равной площади, равной удаленности от материка и расположенные в одной климатической зоне. Отличие только в том, что один из них вулканического происхождения, а другой - биогенного (коралловый). Эти острова отличаются богатством видового состава. Чем определяются эти различия и в чем они заключаются?
4. Даны два острова одного возраста, равной площади, равной удаленности от материка и расположенные в одной климатической зоне. Отличие только в том, что один из них вулканического происхождения, а другой - биогенного (коралловый). Эти острова отличаются богатством видового состава. Чем определяются эти различия и в чем они заключаются?
|
|
|
5. Почему виды, для которых характерна низкая плотность популяций, имеют повышенную предрасположенность к вымиранию в условиях изолятов?
6. Подберите реальные примеры влияния интродукции на сообщества изолятов.
§ 18. Некоторые особенности водных экосистем
Любой водоем: естественный или искусственный, глубокий или мелкий, с пресной или соленой водой, лужа или океан - экосистема. Водная среда обитания имеет особенности, определяющие специфику жизнедеятельности организмов-гидробионтов.
Гидробионты - обитатели водной среды.
Какие химические и физические свойства воды позволяют живым организмам использовать ее как среду обитания?
Классификация водных экосистем.
Чтобы разобраться в многообразии водных экосистем, прибегнем к традиционному способу – классификации, возможные варианты которой представены в таблице справа и на рис. 3.8.
Особые свойства лентических систем.
 Одной из примечательных особенностей лентических систем является сходство видового состава при очевидной изолированности сообществ. Одни и те же виды заселяют озера определенных природных зон в пределах континентов и даже на соседствующих материках. Для объяснения этого странного факта выдвинуто несколько гипотез. Согласно одной из них, лентические экосистемы по эволюционным меркам слишком молоды, и для завершения процессов видообразования прошло еще недостаточно времени.
Одной из примечательных особенностей лентических систем является сходство видового состава при очевидной изолированности сообществ. Одни и те же виды заселяют озера определенных природных зон в пределах континентов и даже на соседствующих материках. Для объяснения этого странного факта выдвинуто несколько гипотез. Согласно одной из них, лентические экосистемы по эволюционным меркам слишком молоды, и для завершения процессов видообразования прошло еще недостаточно времени.
Действительно, большинство крупных озер Северного полушария образовалось не раньше ледникового периода. Исключений очень немного, например, озеро Байкал в Сибири и озеро Виктория в Северной Америке. Именно эти озера отличаются особенным видовым составом и большим количеством эндемичных видов. Согласно другой гипотезе, изоляция лентических экосистем – кажущееся явление. В нарушении изоляции особую роль отводят форезии. Еще одной особенностью лентических экосистем является неравномерность распределения факторов, необходимых для нормальной жизнедеятельности продуцентов, между гиполимнионом и эпилимнионом (см. схему "Классификация водных экосистем").
Эпилимнион богат светом и кислородом, но беден минеральными биогенами, оседающими в гиполимнион. Последнему света и кислорода достается мало. Перемешиванию воды препятствует прогревание поверхностных слоев воды (на дно опускаться будет только холодная вода, имеющая большую плотность). Теплый эпилимнион отделен от холодного гиполимниона зоной термоклина (граница теплой и холодной воды).
|
|
|
Аналогично складываются условия в морских пелагических экосистемах. Возникает парадоксальная ситуация: мелкие заросшие озера с "цветущей" водой более продуктивны, чем глубокие и чистые. В мелких озерах больше рыбы, чем в глубоких (правда, в глубоких она крупнее). Перемешивание слоев и создание благоприятных условий происходит в относительно холодную погоду: в умеренных широтах – весной и осенью, в теплых районах – зимой, в тропиках – постепенно и нерегулярно.
Особенности морских экосистем в зонах апвеллинга.
Глубина океана огромна. Живые организмы встречаются на всех уровнях, но их распределение крайне неравномерно. Объяснить этот факт можно по аналогии с крупными лентическими системами: факторы, необходимые для жизни, рассредоточены в поверхностных и глубинных слоях воды. Этим объясняется резкий всплеск биомассы литоральных и сублиторальных экосистем, меньшая населенность эпипелагиальных и абиссальных экосистем и малочисленные организмы батипелагиальной зоны. Однако в некоторых областях океана, именуемых областями апвеллинга, картина сказочным образом меняется:
- преобладает пелагическое сообщество,
- огромное количество планктона,
- колоссальные популяции рыб,
- короткие цепи питания (многие хищные виды ракообразных и рыб переходят на растительную пищу),
 - над морем кружатся огромные стаи птиц,
- над морем кружатся огромные стаи птиц,
- на дно оседает очень много органики, что способствует бурному развитию аллохтонных бентосных экосистем. В противоположность этой идиллической картине морского сообщества прилегающие к зонам апвеллинга районы суши пустынны, и только по береговой кромке туманы поддерживают чахлую растительность.
Чем же можно объяснить подобные особенности? Для ответа проанализируем расположение пяти главных зон прибрежных апвеллингов:
Рис. 3.9. Области апвеллинга
1) юго-западное побережье Северной Америки,
2) северо-западное побережье Южной Америки,
3) северо-западное побережье Африки,
4) юго-западное побережье Африки,
5) южное побережье Аравийского полуострова.
Обратите внимание: все крупные апвеллинги располагаются у западных побережий близких к экватору территорий с крутыми материковыми склонами. Вращением Земли определяется преобладающее направление ветров в этих районах: с востока на запад, т.е. с материка в океан. Ветер сухой, т.к. несет воздушные массы с материка, и поэтому прилегающие к побережью территории пустыни. Отгоняя от берега поверхностные слои воды, ветровые потоки вызывают активное перемешивание, и биогены выносятся на поверхность. Создаются благоприятные условия для бурного развития планктона и апвеллинговых экосистем. Периодически в зонах апвелинга возникают очень неприятные явления: при изменении направления ветра перемешивание прекращается, и в подповерхностных слоях эпипелагиали, богатых биогенами, но бедных кислородом, происходит активное размножение анаэробных одноклеточных растительных жгутиконосцев динофлагеллат. Клетки этих организмов окрашены в красный цвет, продукты их обмена веществ ядовиты. Подобное явление носит название "красные приливы" и приводит к массовой гибели рыбы. В это время категорически запрещена рыбная ловля, т.к. использование в пищу отравленной рыбы может привести к гибели людей.
Краткое содержание § 18
Водная среда обитания имеет особенности, определяющие специфику жизнедеятельности организмов-гидробионтов. Принято выделять пресноводные и морские экосистемы. Экосистемы пресных вод разделяют на лентические (стоячие воды), лотические (текучие воды), заболоченные земли и болота. Среди морских экосистем выделяют бенталь (придонные экосистемы) и пелагиаль (экосистемы водных толщ). Особенностью лентических и морских экосистем является неравномерность распределения факторов, необходимых для нормальной жизнедеятельности продуцентов, между поверхностными и глубокими слоями воды. Поверхностные воды богаты светом и кислородом, но бедны минеральными биогенами, оседающими вниз. Глубоким слоям воды света и кислорода достается мало. Перемешиванию препятствует прогревание поверхностных слоев - на дно опускается только холодная вода, имеющая большую плотность. Перемешивание слоев и создание благоприятных условий происходит в относительно холодную погоду и(или) под воздействием ветров. В некоторых участках океана, которые называются областями апвелинга, ветровые потоки вызывают активное перемешивание, и биогены выносятся на поверхность. Создаются благоприятные условия для бурного развития планктона и апвеллинговых экосистем.
Задания.
1. Сравните экосистемы: 1) сублиторали и абиссали, 2) эпипелагиали и батипелагиали, 3) эпипелагиали и абиссали,
используя следующие параметры: а) наиболее характерный тип цепей питания, б) автохтонность или аллохтонность экосистем, в) структура пирамиды биомасс, г) наиболее вероятные типы сукцессий, д) условия, необходимые для достижения климакса.
Результаты сравнения представьте в виде таблицы.
2. Рыбаки выбирают место для рыбалки. Один предлагает остановиться у заросшего "цветущего" пруда, другой предлагает пройти еще немного до следующего пруда с чистой водой и прекрасными песчаными пляжами.
Какой пруд им следует выбрать, если основная задача - наловить побольше рыбы?
 3. Вы - эксперт-эколог строительной организации. Вам предлагается оценить последствия строительства плотины для микроклимата и состояния водных экосистем района водохранилища и территории ниже по течению реки.
3. Вы - эксперт-эколог строительной организации. Вам предлагается оценить последствия строительства плотины для микроклимата и состояния водных экосистем района водохранилища и территории ниже по течению реки.
Имеются два проекта (рис. 3.10): в одном слив воды из водохранилища устроен в нижней части плотины ("нижний слив"), а в другом - организован слив поверхностной воды ("верхний слив"). Вынесите заключение и аргументируйте его.
4. При искусственном орошении пустынных земель, прилегающих к зонам апвеллинга, получают очень высокие урожаи. Чем можно объяснить высокое потенциальное плодородие этих территорий?
|
|
|
