 |


Формирование «систем» дифференцировочных условных рефлексов
|
|
|
|
В работах Л. Г. Воронина (1984) также было показано, что при комбинации отдельных УР возникают новые свойства, не сводимые к свойствам суммы исходных рефлексов. Происходит образование так называемых систем дифференцировочных УР, или просто «систем». Такие системы Л. Г. Воронин считал целостными функциональными единицами, из которых складывается психическая деятельность.
У животного вырабатывают несколько дифференцировочных УР, объединенных в систему таким образом, что выполнение одного УР зависит от усвоения другого.
Одна из таких процедур составляет необходимый этап в изучении способности к транзитивному заключению — форме дедуктивного мышления, свойственной и животным, и человеку.
Программа эксперимента по выработке систем дифференцировок состоит в следующем. Животное учится дифференцировать несколько пар раздражителей (как правило, цветовых стимулов). При выработке дифференцировки каждой следующей пары стимулов производится смена сигнального значения части из них в соответствии с определенным правилом: стимул, бывший положительным в паре А— В+ (где «В+» — подкрепляемый стимул, а «А—» — неподкрепляемый), становится отрицательным в паре В— С+. В процессе обучения животное должно усвоить информацию о том, что значения стимулов В, С, D могут быть как положительными (В+, С+, D+), так и отрицательными (В—, С—,
D—), в зависимости от пары, в которой они в данный момент предъявляются. Стимулы А- и F+, т.е. первый и последний в последовательности, остаются во всех комбинациях только положительным (F) или только отрицательным (А). Высокая доля правильных выборов при чередовании всех пар стимулов, когда животное выбирает, например, стимул С в паре В— С+, но не реагирует на него в паре С— D+, свидетельствует, что оно усвоило принцип данной системы. Такую систему дифференцировок успешно усваивают дети, начиная с 4 лет (Bryant, Trabasso, 1971), взрослые шимпанзе (Gillan, 1981; Boysen, Berntson, 1995), а также голуби и вороны (Зорина и др., 1989). Межвидовые различия в динамике и успешности формирования такой системы дифференцировок у этих видов птиц оказались незначительными и недостоверными. В то же время при обратимых последовательных переделках и формировании установки на обучение голуби существенно отставали как от врановых, так и от приматов.
|
|
|
В0 Метод выработки дифференцировочных УР в разных его комбинациях является одним из ведущих в физиологии ВНД и ис-в пользуется как инструмент для исследования целого ряда проблем.
К этим проблемам, помимо оценки возможностей органов чувств животных разных видов, относится изучение следующих более сложных когнитивных способностей:
* к обобщению;
* к транзитивному заключению;
* к формированию установки на обучение.
Такие экспериментальные исследования будут подробнее рассмотрены ниже.
3.4. Когнитивные (познавательные) процессы 3.4.1. Общая характеристика
Термин «.когнитивные», или и познавательные», процессы употребляют для обозначения тех видов поведения животных и человека, в основе которых лежит не условнорефлекторный ответ на воздействие внешних стимулов, а формирование внутренних (мысленных) представлений о событиях и связях между ними.
И. С. Бериташвили называл их психонервными образами, или психонервными представлениями, Е. Н. Соколов — нервной моделью стимула, Л. А. Фирсов (1972; 1993) и Т. А. Натишвили (1987) — образной памятью. Д. Мак-Фарленд (1988) подчеркивает, что когнитивная деятельность животных относится к мыслительным процессам, которые
ячастую недоступны прямому наблюдению, однако их существование возможно выявить в эксперименте.
|
|
|
Наличие представлений обнаруживается в тех случаях, когда субъект (человек или животное) совершает действие без влияния какого бы то ни было физически реального стимула. Такое возможно, например, когда он извлекает информацию из памяти или мысленно восполняет отсутствующие элементы действующего стимула. В то же время формирование мысленных представлений может никак не проявляться в исполнительной деятельности организма и обнаружится лишь позднее, в какой-то определенный момент.
Внутренние представления могут отражать самые разные типы сенсорной информации, не только абсолютные, но и относительные признаки стимулов, а также соотношения между разными стимулами и между событиями прошлого опыта. По образному выражению, животное создает некую внутреннюю картину мира, включающую комплекс представлений «что», «где», «когда». Они лежат в основе обработки информации о временных, числовых и пространственных характеристиках среды и тесно связаны с процессами памяти. Различают также образные и абстрактные (отвлеченные) представления (Premack, 1983). Последние рассматривают как основу формирования довербаль-ных понятий (см. гл. 5).
Метод отсроченных реакций. Мысль о существовании у животных некоего «процесса представления», т.е. такой активности мозга, которая соответствует полученной ранее стимуляции, но которая может поддерживаться в ее отсутствие, была впервые высказана У.Хантером в 1913 г. Для оценки способности животного реагировать на воспоминание о стимуле в отсутствие этого реального стимула У. Хантер предложил метод отсроченных реакций (2.4.4).
Животное (в опытах Хантера — енота) помещали в клетку с тремя одинаковыми и симметрично расположенными дверцами для выхода. Над одной из них на короткое время зажигали лампочку, а потом еноту давали возможность подойти к любои из дверец. Если он выбирал дверцу, над которой зажигалась лампочка, то получал подкрепление. При соответствующей тренировке животные выбирали нужную дверцу даже после 25-секундной отсрочки — интервала между выключением лампочки и возможностью сделать выбор.
В опытах других исследователе!) задача ставится иначе. На глазах у голодного животного в один из двух (или трех) ящиков помещают корм. По истечении периода отсрочки животное выпускают из клетки или (как на рис. 3.9) убирают отделяющую его преграду. Его задача выбрать ящик с кормом.
|
|
|
Выполнение теста на отсроченные реакции на неслучайном уровне считается доказательством наличия у животного мысленного представления о спрятанном предмете (его образа), т.е. существования какой-то активности мозга, которая в этом случае подменяет информацию от органов чувств. С помощью этого метода были исследованы представители различных видов животных и было подтверждено, что их поведение может направляться не только действующими в данный момент
стимулами, но также и хранящимися в памяти следами, образами или представлениями об отсутствующих стимулах (рис. 3.9).
Опыты на разных животных были дополнены исследованиями на детях. Эту методику стали довольно широко применять и в нашей стране, начиная с 30-х годов, но многие ученики И. П. Павлова (например, Э. Г. Вацуро) отождествляли ее с методом следового УР, когда выполнение условной реакции происходит как бы на основе «следов действия УС». Метод отсроченных реакций стали использовать как тест на форму особой (образной или декларативной) памяти, которая не идентична памяти, формирующейся при УР (см. словарь терминов).
Между условнорефлекторной (ассоциативной или процедурной) и образной (декларативной) памятью существуют тонкие функциональные различия. От особенностей их взаимодействия зависят, например, различия в организации сложного поведения низших и высших обезьян (Фирсов, 1972; 1993).
В классическом тесте на отсроченные реакции виды животных проявляют себя по-разному. Собаки, например, после того как корм положен в один из ящиков, ориентируют тело по направлению к нему и сохраняют эту неподвижную позу в течение всего периода отсрочки, а по ее окончании сразу бросаются вперед и выбирают нужный ящик. Другие животные в подобных случаях не сохраняют определенной позы и могут даже разгуливать по клетке, что не мешает им, тем не менее, правильно обнаруживать приманку. У шимпанзе формируется не просто представление об ожидаемом подкреплении, но ожидание определенного его вида. Так, если вместо показанного в начале опыта банана после отсрочки обезьяны обнаруживали салат (менее ими любимый), то отказывались его брать и искали банан. Мысленные представления контролируют и гораздо более сложные формы поведения. Многочисленные свидетельства этого были получены и в специальных экспериментах, и в наблюдениях за повседневным поведением обезьян в неволе и естественной среде обитания (см., например: Ладыгина-Коте, 1923; 1935; Гудолл, 1992; подробнее об этом см. гл. 7).
|
|
|
Тест на оценку представлений о константности свойств предметов. Один из способов изучения роли представлений в поведении и психике животного — тест на «константность свойств предмета» («object permanence»), т.е. на способность субъекта понимать, что предмет, скрывшийся из поля зрения, продолжает существовать, может быть отыскан и не меняет при этом своих свойств. Тест широко применяется в исследованиях психического развития животных и человека (Пиаже, 1969; Бауэр, 1979).
Для оценки способности животных оперировать представлениями о константности свойств предмета существуют тесты:
С
• на «неисчезаемость»;
• на «вмещаемость»;
• на «перемещаемость» и др.
Термины были введены Л. В. Крушин-ским (1986) и соответствующие тесты названы им «эмпирическими законами». Они соответствует 4-й и 6-й стадиям развития по Пиаже (1969) (подробнее об оценке этих способностей у животных см. 4.6.1).
f\ В основе понимания животным или ребенком принципа «неисчезае-мости» лежит эмпирическое знание того, что предметы существуют, даже если в данный момент они недоступны непосредственному восприятию органами чувств.
По мнению Л. В. Крушинского, понимание животным этого принципа составляет необходимое условие для проявления способности к экстраполяции направления движения стимула и решению ряда других элементарных логических задач (см. гл. 4).
«Мысленный план» лабиринта. Одним из первых гипотезу о роли представлений в обучении животных выдвинул Э. Толмен в 30-х годах XX века (1930, 1997). Исследуя поведение крыс в лабиринтах разной конструкции, он пришел к выводу, что общепринятая в то время схема «стимул-реакция» не может удовлетворительно описать поведение животного, усвоившего ориентацию в такой сложной среде, как лабиринт. Толмен высказал предположение, что в период между действием стимула и ответной реакцией в мозге совершается определенная цепь процессов («внутренние или промежуточные переменные»), которые определяют последующее поведение. Сами эти процессы, по мнению Толмена, можно исследовать строго объективно по их функциональному проявлению в поведении.
|
|
|

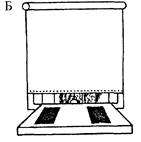

Рис. 3.9. Схема опытов по изучению отсроченных реакций.
А — демонстрация приманки и пустого контейнера; Б — приманку и пустой контейнер закрывают от животного непрозрачным экраном; В — экран убирают, и животное имеет возможность достать приманку, положение которой не изменилось При следующем предъявлении этого теста контейнер с приманкой может располагаться слева.
В процессе обучения у животного формируется «когнитивная карта» всех признаков лабиринта, или его «мысленный план». Затем на основе этого «плана» животное выстраивает свое поведение.
Толмен и его последователи исследовали формирование «мысленного плана» («карты») с помощью лабиринтов разных конструкций, траекторию движения в которых животные могли менять в зависимости от того, были ли им доступны более короткие пути.
В работе Н. Чапиуса и П. Скардигли (цит. по: Реагсе, 1998) хомячка помещали в камеру А гексагонального лабиринта (рис. 3.10А), в котором ему «надлежало» пройти за пищей по пути, указанному штриховой линией. От этой дороги хомячок не отклонялся, поскольку все входы в другие отсеки были закрыты. После запоминания этого пути во время целого ряда предъявлении задачи в лабиринте открывали все дверцы, за исключением той, которая позволяла попасть к корму по прямой. Теперь кратчайшим путем к пище стала траектория через центр (сплошная линия). Этот путь можно уверенно найти, только имея «мысленный план» данного лабиринта. После определенной тренировки хомячки выбирали этот путь достоверно чаще, чем остальные.
Два других примера, взятых из работы самого Толмсна, показывают, как животное находит новый путь к пище, если знакомая дорога перекрыта (рис. 3.10Б и В).
Образование «мысленного плана» может происходить и в отсутствие подкрепления, в процессе ориентировочно-исследовательской активности. Этот феномен Толмен назвал латентным обучением (см. 3.4.2).
Сходных взглядов на организацию поведения придерживался И. С. Бериташвили (1974). Ему принадлежит термин — «поведение, направляемое образом». Бериташвили продемонстрировал способность собак к формированию представлений о структуре пространства, а также «психонервных образов» предметов. Ученики и последователи И. С. Бериташвили показали пути видоизменения и совершенствования образной памяти в процессе эволюции (Натишвили, 1987), а также в онтогенезе (Бадридзе, 1987), базируясь на данных по пространственной ориентации животных.
Гипотеза о когнитивных механизмах многих форм поведения закономерно возникала у исследователей, пытавшихся осмыслить данные самых разнообразных экспериментов. Американский ученый Д. Гриф-фин (Griffin, 1984) в книге «Мышление животных» («Animal thinking») в популярной форме рассматривает факты, хотя и не всегда действительно относящиеся к сфере мышления животных, но явно не получавшие объяснения в терминах бихевиоризма (например, орудийная деятельность и сложные виды коммуникации).
Когнитивная психология человека — направление, возникшее как реакция на господство идей бихевиоризма в области изучения психики человека, — оказала влияние на исследования когнитивных процессов
 Рис. 3.10. Лабиринты, в которых исследовали формирование «мысленных карт» у животных.
Рис. 3.10. Лабиринты, в которых исследовали формирование «мысленных карт» у животных.
А —гексагональный лабиринт; Б — усвоение общего плана строения лабиринта при изменении его конфигурации. Слева — голодная крыса обучается находить путь к пище, следуя по траектории а-б-в-г-д-е. После упрочения навыка животное помещают в точку а, но теперь из круглой камеры можно выйти по множеству рукавов за исключением одного (в). При наличии у животного «мысленного плана» оно чаще, чем случайно, выходит к пище по рукаву 5. В — крысу помещали в точку старта (с) и обучили находить корм (К). Постепенно у нее сформировался «мысленный план» лабиринта. После того как убрали перегородку (П), она стала бегать по более короткому пути, который обозначен пунктирной линией.
у животных. Проведение аналогий между работой мозга и вычислительной машиной позволило описывать память животных как процесс переработки информации с привлечением соответствующего математического аппарата и понятий теории информации (Солсо, 1996).
Одно из наиболее популярных направлений в анализе когнитивных процессов у животных — это анализ обучения «пространственным» навыкам (главным образом, благодаря введению в лабораторную практику методов водного и радиального лабиринтов, см. 3.4.2). Существует даже тенденция считать эти феномены основным проявлением
когнитивной деятельности животных (в действительности же в эту категорию входит значительно более широкий круг индивидуально-приспособительных реакций животных, в том числе довербальные понятия, способность к усвоению и использованию символов, которая была обнаружена при обучении языкам-посредникам, и др.).
Понятие, «когнитивная деятельность животных» включает способность к разным видам обучения и элементы довербального мышления.
На формировании представлений основаны следующие виды обучения животных:
* латентное обучение;
* пространственное обучение;
* выбор по образцу;
* заучивание последовательностей.
Латентное обучение
По определению У. Торпа, латентное обучение — это «...образование связи между индифферентными стимулами или ситуациями в отсутствие явного подкрепления».
Элементы латентного обучения присутствуют практически в любом процессе обучения, но могут быть выявлены только в специальных опытах.
В естественных условиях латентное обучение возможно благодаря исследовательской активности животного в новой ситуации. Оно обнаружено не только у позвоночных. Эту или сходную способность для ориентации на местности используют, например, многие насекомые. Особенно хорошо латентное обучение изучено у перепончатокрылых. Так, пчела или оса, прежде чем улететь от гнезда, совершает «рекогносцировочный» полет над ним, что позволяет ей фиксировать в памяти «мысленный план» данного участка местности.
Наличие такого «латентного знания» выражается в том, что животное, которому предварительно дали ознакомиться с обстановкой опыта, обучается быстрее, чем контрольное, не имевшее такой возможности.
В настоящее время термин «латентное обучение» употребляется редко и лишь в определенном контексте. Однако на самом деле этот феномен достаточно широко распространен. Например, в разделе 4.8.2 будет показано, что оно сопровождает выработку простых инструментальных УР (открывание кормушек разного цвета). Птицы (вороны и голуби) запоминают число единиц подкрепления, получаемого при действии стимулов разного цвета, а затем используют эту информацию в новой ситуации (Зорина и др., 1991). В гл. 5 мы покажем, что феномен, который Л. А.Фирсов называет «вторичным обучением», по существу можно считать одним из вариантов латентного обучения.
|
|
|
