 |


Основные генетические процессы в клетках
|
|
|
|
Клеточный цикл и его регуляция
Клеточный цикл - это период жизни клетки от одного деления до другого или от деления до смерти. Клеточный цикл состоит из интерфазы (период вне деления) и самого клеточного деления.
Если клетка собирается когда-нибудь делиться, то интерфаза будет состоять из трех периодов. Сразу после выхода из митоза клетка вступает в пресинтетический или G1 период, далее переходит в синтетический или S период и потом - в постсинтетический или G2 период. G2 периодом заканчивается интерфаза и после нее клетка вступает в следующий митоз.
Если клетка не планирует снова делиться, то она как бы выходит из клеточного цикла и вступает в период покоя, или G0 период. Если клетка, находящаяся в G0 периоде, снова захочет делиться, то она выходит из G0 периода и вступает в G1 период.
Длительность периодов клеточного цикла различна. Наибольшим постоянством отличаются S, G2 периоды и митоз, а G1 период очень вариабелен. Так, G1 период может продолжаться от 2-4 ч до нескольких недель или даже месяцев. Как правило, продолжительность S-периода варьирует от 6 до 8 ч, а G2 периода - от нескольких часов до получаса. Длительность митоза составляет в среднем от 40 до 90 минут. Причем самой короткой фазой митоза можно считать анафазу. Она занимает всего несколько минут.
G1 период характеризуется высокой синтетической активностью, которая должна увеличить свой обьем до размера материнской клетки, а значит, и количество органелл, различных веществ.Непонятно почему, но клетка, прежде чем вступить в следующий митоз, должна иметь размер, равный материнской клетке. И пока этого не произойдет, клетка продолжает оставаться в G1 периоде. Видимо, единственным исключением из этого является дробление, при котором бластомеры делятся, не достигая размеров исходных клеток.
|
|
|
В конце G1 периода принято различать специальный момент, называемый R-точкой (точка рестрикции, R-пункт), после которого клетка обязательно в течение нескольких часов (обычно 1-2) вступает в S период. Период времени между R-точкой и началом S периода можно рассматривать в качестве подготовительного для перехода в S период.
Самый главный процесс, который идет в S периоде, - это удвоение или редупликация ДНК. Все остальные реакции, происходящие в это время в клетке, направлены на обеспечение синтеза ДНК. К таким вспомогательным процессам можно отнести синтез гистоновых белков, синтез ферментов, регулирующих и обеспечивающих синтез нуклеотидов и образование новых нитей ДНК.
Сущность G2 периода не совсем понятна в настоящее время, однако в этот период происходит образование веществ, необходимых для самого процесса митоза. В G2 периоде происходит синтез белков, из которых образуются микротрубочки веретена деления (тубулин, динеин, нексин, спектрин), происходит синтез АТФ.
Сейчас является установленным, что прохождение клетки по всем периодам клеточного цикла строго контролируется. При движении клеток по клеточному циклу в них появляются и исчезают, активируются и ингибируются специальные регуляторные молекулы, которые обеспечивают:
1) прохождение клетки по определенному периоду клеточного цикла
2) переход из одного периода в другой.
Причем прохождение по каждому периоду, а также переход из одного периода в другой контролируется различными веществами. Сейчас мы попробуем выяснить, что же это за вещества и что они делают.
Общая ситуация выгладит так. В клетке постоянно присутствуют специальные белки-ферменты, которые путем фосфорилирования других белков регулируют активность генов, ответственных за прохождение клетки по тому или иному периоду клеточного цикла. Эти белки-ферменты называются циклин-зависимыми протеинкиназами (cdc). Они присутствуют в клетке постоянно, независимо от периода клеточного цикла, то есть они имеются в избытке. Другими словами, их синтез или количество не лимитирует или не регулирует прохождение клеток по клеточному циклу. Однако при патологии, если синтез их нарушен, снижено их количество или имеются мутантные формы с измененными свойствами, то это, конечно же, может повлиять на течение клеточного цикла.
|
|
|
Почему же такие циклин-зависимые протеинкиназы сами не могут регулировать прохождение клеток по периодам клеточного цикла. Оказывается, что они находятся в клетках в неактивном состоянии, а для того чтобы они активировались и начали работать, необходимы специальные активаторы. Ими являются циклины. Их также много разных типов, но они присутствуют в клетках не постоянно: то появляются, то исчезают. Появление и исчезновение циклинов обусловлено их синтезом и быстрым разрушением, то есть наличие циклинов лимитирует или регулирует работу циклин-зависимых протеинкиназ. Причем синтез каждого циклина происходит в строго определенный период клеточного цикла. В один период образуются одни циклины, а в другой - другие. Так, например, прохождение клетки по G1 периоду клеточного цикла обеспечивает комплекс циклин-зависимой протеинкиназы-2 (cdk2) и циклина D1, циклин-зависимой протеинкиназы-5 (cdk5) и циклина D3. Прохождение через специальную точку рестрикции (R-пункт) периода G1 контролирует комплекс cdc2 и циклина С. Переход клетки из G1 периода клеточного цикла в S период контролирует комплекс cdk2 и циклиа Е. Для перехода клетки из S периода в G2 период необходим комплекс cdk2 и циклин А. Циклин-зависимая протеинкиназа-2 (cdc2) и циклин В участвуют в переходе клетки из G2 периода в митоз (М период). Циклин H в соединении с cdk7 необходим для фосфорилирования и активациии cdc2 в комплексе с циклином В.
Регуляция клеточного цикла
G1 период cdk2 + циклин D1 cdk5 + циклин D3
R-пункт периода G1 cdc2 + циклин С
переход из G1 в S период cdk2 + циклин Е
переход из S в G2 период cdk2 + циклин А
переход из G2 периода в митоз (М период) cdc2 + циклин В
циклин H + cdk7 необходим для фосфорилирования и активациии cdc2 в комплексе с циклином В
|
|
|
Апоптоз
Апоптоз представляет собой вариант клеточной смерти, которая имеет место в нормальных физиологических условиях и сама клетка является активным участником своей собственной смерти. Апоптоз наиболее часто наблюдается в течение обычного клеточного обновления, при поддержании тканевого гомеостаза, в эмбриогенезе, при индукции и поддержании иммунологической толерантности, развитии нервной системы, тканевой атрофии.
Сейчас является общепризнанным, что большинство, но не все, физиологических смертей клеток происходит путем апоптоза. Клетки, вступающие в апоптоз, имеют специфические морфологические и биохимические характеристики. При апоптозе происходит смерть отдельных клеток, а не какой-то группы. Апоптоз индуцируется физиологическими стимулами.
Биохимическими маркерами апоптоза являются деградация геномной ДНК. Это необратимое событие и направляет клетку к гибели и проявляется до наступления изменений в клеточной проницаемости. В большинстве клеток такую фрагментацию ДНК вызывают кальций и магний-зависимые ядерные эндонуклеазы, которые селективно разрезают ДНК на сайты, локализованные между нуклеосомами (линкерные участки ДНК), и приводят к образованию моно- и олиго-нуклеосомных фрагментов ДНК. Процесс активации ферментов тонко регулируется и является АТФ-зависимым.
Процесс апоптоза регулируется специальными генами. Усиление работы генов Cip1 (p21), Bax (p21), Daax, FAF-1, FADD, TRADD, RAIDD, RIP, SIVA, FLIP, CAS, TIA-1/TIAR, TDAG51 и др., белков p53, р21 ускоряет течение апоптоза. Активация мембранного рецепторного гликопротеина, кодируемого геном Fas/Apo-1, приводит к апоптотической гибели лимфоидных клеток, экспрессирующих этот рецептор. Продукты генов семейства Bcl-2, Bad, Bag1 препятствуют апоптозу. В клетках с поврежденной ДНК апоптозу предшествует экспрессия протоонкогенов fos, myc и p53. Апоптоз иногда бывает подавленым. Например, при аутоиммунном лимфопролиферативном синдроме наблюдается угнетенте апоптоза лимфоцитов вследствие мутации гена, кодирующего гликопротеин Fas/Apo-1.
|
|
|
Митоз
Различают следующие четыре фазы митоза: профаза, метафаза, анафаза и телофаза. В профазе хорошо видны центриоли — образования, находящиеся в клеточном центре и играющие роль в делении дочерних хромосом животных. (Напомним, что у высших растений нет центриолей в клеточном центре, который организует деление хромосом). Мы же рассмотрим митоз на примере животной клетки, поскольку присутствие центриоли делает процесс деления хромосом более наглядным. Центриоли делятся и расходятся к разным полюсам клетки. От центриолей протягиваются микротрубочки, образующие нити веретена деления, которое регулирует расхождение хромосом к полюсам делящейся клетки.
В конце профазы ядерная оболочка распадается, ядрышко постепенно исчезает, хромосомы спирализуются и в результате этого укорачиваются и утолщаются, и их уже можно наблюдать в световой микроскоп. Еще лучше они видны на следующей стадии митоза — метафазе.
В метафазе хромосомы располагаются в экваториальной плоскости клетки. При этом хорошо видно, что каждая хромосома, состоящая из двух хроматид, имеет перетяжку — центромеру. Хромосомы своими центромерами прикрепляются у нити веретена деления. После деления центромеры каждая хроматида становится самостоятельной дочерней хромосомой.
Затем наступает следующая стадия митоза — анафаза, во время которой дочерние хромосомы (хроматиды одной хромосомы) расходятся к разным полюсам клетки.
Следующая стадия деления клетки — телофаза. Она начинается после того, как дочерние хромосомы, состоящие из одной хроматиды, достигли полюсов клетки. На этой стадии хромосомы вновь деспирализуются и приобретают такой же вид, какой они имели до начала деления клетки в интерфазе (длинные тонкие нити). Вокруг них возникает ядерная оболочка, а в ядре формируется ядрышко, в котором синтезируются рибосомы. В процессе деления цитоплазмы все органоиды (митохондрии, комплекс Гольджи, рибосомы и др.) распределяются между дочерними клетками более или менее равномерно.
Таким образом, в результате митоза из одной клетки получаются две, каждая из которых имеет характерное для данного вида организма число и форму хромосом, а следовательно, постоянное количество ДНК.
Весь процесс митоза занимает в среднем 1-2 ч. Продолжительность его несколько различна для разных видов клеток. Зависит он также от условий внешней среды (температуры, светового режима и других показателей).
Биологическое значение митоза заключается в том, что он обеспечивает постоянство числа хромосом во всех клетках организма. Все соматические клетки образуются в результате митотического деления, что обеспечивает рост организма. В процессе митоза происходит распределение веществ хромосом материнской клетки строго поровну между возникающими из нее двумя дочерними клетками. В результате митоза все клетки организма получают одну и ту же генетическую информацию.
|
|
|
Мейоз
Половое размножение животных, растений и грибов связано с формированием специализированных половых клеток.
Мейоз — особый тип деления клеток, в результате которого образуются половые клетки.
В отличии от митоза, при котором сохраняется число хромосом, получаемых дочерними клетками, при мейозе число хромосом в дочерних клетках уменьшается вдвое.
Процесс мейоза состоит из двух последовательных клеточных делений — мейоза I (первое деление) и мейоза II (второе деление).
Удвоение ДНК и хромосом происходит только перед мейозом I.
В результате первого деления мейоза, называемого редукционным, образуются клетки с уменьшенным вдвое числом хромосом. Второе деление мейоза заканчивается образованием половых клеток. Таким образом, все соматические клетки организма содержат двойной, диплоидный (2n), набор хромосом, где каждая хромосома имеет парную, гомологичную хромосому. Зрелые половые клетки имеют лишь одинарный, гаплоидный (n), набор хромосом и соответственно вдвое меньшее количество ДНК.
Фазы мейоза
Во время профазы I мейоза двойные хромосомы хорошо видны в световой микроскоп. Каждая хромосомы состоит из двух хроматид, которые связаны вместе одной центромерой. В процессе спирализации двойные хромосомы укорачиваются. Гомологичные хромосомы тесно соединяются друг с другом продольно (хроматида к хроматиде), или, как говорят, конъюгируют. При этом хроматиды нередко перекрещиваются или перекручиваются одна вокруг другой. Затем гомологичные двойные хромосомы начинают как бы отталкиваться друг от друга. В местах перекреста хроматид происходят поперечные разрыва и обмены их участками. Это явление называют перекрестом хромосом. Одновременно, как и при митозе, распадется ядерная оболочка, исчезает ядрышко, образуются нити веретена. Отличие профазы I мейоза от профазы митоза состоит в конъюгации гомологичных хромосом и взаимном обмене участками в процессе перекреста хромосом.
Характерный признак метафазы I — расположение в экваториальной плоскости клетки гомологичных хромосом, лежащих парами. Вслед за этим наступает анафаза I, во время которой целые гомологичные хромосомы, каждая состоящая из двух хроматид, отходят к противоположным полюсам клетки. Очень важно подчеркнуть одну особенность расхождения хромосом на этой стадии мейоза: гомологичные хромосомы каждой пары расходятся в стороны случайным образом, независимо от хромосом других пар. У каждого полюса оказывается вдвое меньше хромосом, чем было в клетке при начале деления. Затем наступает телофаза I, во время которой образуются две клетки с уменьшенным вдвое числом хромосом.
Интерфаза короткая, так как синтеза ДНК не происходит. Далее следует второе мейотическое деление (мейоз II). Оно отличается от митоза только тем, что количество хромосом в метафазе II вдвое меньше, чем количество хромосом в метафазе митоза у того же организма. Поскольку каждая хромосома состоит из двух хроматид, то в метафазе II центромеры хромосом делятся, и к полюсам расходятся хроматиды, которые становятся дочерними хромосомами. Только теперь наступает настоящая интерфаза. Из каждой исходной клетки возникают четыре клетки с гаплоидным набором хромосом.
Разнообразие гамет
Рассмотри мейоз клетки, имеющей три пары хромосом (2n = 6). В этом случае после двух мейотических делений образуются четыре клетки с гаплоидным набором хромосом (n = 3). Поскольку хромосомы каждой пары расходятся в дочерние клетки независимо от хромосом других пар, равновероятно образование восьми тиров гамет с различным сочетанием хромосом, присутствовавших в исходной материнской клетке.
Еще большее разнообразие гамет обеспечивается конъюгацией и перекрестом гомологичных хромосом в профазе мейоза, что имеет очень большое общебиологическое значение.
Биологическое значение мейоза
Если бы в процессе мейоза не происходило уменьшения числа хромосом, то в каждом следующем поколении при слиянии ядер яйцеклетки и сперматозоида число хромосом увеличивалось бы бесконечно. Благодаря мейозу зрелые половые клетки получают гаплоидное (n) число хромосом, при оплодотворении же восстанавливается свойственное данному виду диплоидное (2n) число. При мейозе гомологичные хромосомы попадают в разные половые клетки, а при оплодотворении парность гомологичных хромосом восстанавливается. Следовательно, обеспечивается постоянных для каждого вида полных диплоидный набор хромосом и постоянное количество ДНК.
Происходящие в мейозе перекрест хромосом, обмен участками, а также независимое расхождение каждой пары гомологичных хромосом определяют закономерности наследственной передачи признака от родителей потомству. Из каждой пары двух гомологичных хромосом (материнской и отцовской), входивших в хромосомный набор диплоидных организмов, в гаплоидном наборе яйцеклетки или сперматозоида содержится лишь одна хромосома. Она может быть:
отцовской хромосомой;
материнской хромосомой;
отцовской с участком материнской;
материнской с участком отцовской.
Эти процессы возникновения большого количества качественно различных половых клеток способствуют наследственной изменчивости.
В отдельных случаях вследствие нарушения процесса мейоза, при нерасхождении гомологичных хромосом, половые клетки могут не иметь гомологичной хромосомы или, наоборот, иметь обе гомологичные хромосомы. Это приводит к тяжелым нарушениям в развитии организма или к его гибели.
Транскрипция и сплайсинг
Транскрипция - первая стадия реализации генетической информации в клетке. В ходе процесса образуются молекулы мРНК, служащие матрицей для синтеза белков, а также транспортные, рибосомальные и другие виды молекул РНК, выполняющие структурные, адапторные и каталитические функции (рис. 4-26).

Рис. 4-26. Схема реализации генетической информации в фенотипические признаки. Реализацию потока информации в клетке можно представить схемой ДНК-"РНК-"белок. ДНК-"РНК обозначает биосинтез молекул РНК (транскрипцию); РНК-"белок означает биосинтез полипептидных цепей (трансляцию).
Транскрипция у эукариотов происходит в ядре. В основе механизма транскрипции лежит тот же структурный.принцип комплементарного спаривания оснований в молекуле РНК (G ≡ C, A=U и Т=А). ДНК служит только матрицей и в ходе транскрипции не изменяется. Синтез молекул РНК начинается в определённых последовательностях (сайтах) ДНК, которые называют промоторы, и завершается в терминирующих участках (сайты терминации). Участок ДНК, ограниченный промотором и сайтом терминации, представляет собой единицу транскрипции - транскриптон. У эукариотов в состав транскриптона, как правило, входит один ген (рис. 4-27), у прокариотов несколько. В каждом транскриптоне присутствует неинформативная зона; она содержит специфические последовательности нуклеотидов.
Соседние транскриптоны могут быть отделены друг от друга нетранскрибируемыми участками ДНК. Разделение ДНК на множество транскриптонов позволяет осуществлять с разной активностью индивидуальное считывание (транскрипцию) разных генов.
В каждом транскриптоне транскрибируется только одна из двух цепей ДНК, которая называется матричной, вторая, комплементарная ей цепь, называется кодирующей. Синтез цепи РНК идёт от 5'- к З'-концу, при этом матричная цепь ДНК всегда антипараллельна синтезируемой нуклеиновой кислоте (рис. 4-28).
Транскрипция не связана с фазами клеточного цикла; она может ускоряться и замедляться в зависимости от потребности клетки или организма в определённом белке.
А. Стадии транскрипции
В процессе транскрипции различают 3 стадии: инициацию, элонгацию и терминацию.
Инициация
Активация промотора происходит с помощью большого белка - ТАТА-фактора, называемого так потому, что он взаимодействует со специфической последовательностью нуклеотидов промотора - ТАТААА- (ТАТА-бокс) (рис. 4-29).
Присоединение ТАТА-фактора облегчает взаимодействие промотора с РНК-полимеразой. Факторы инициации вызывают изменение кон-формации РНК-полимеразы и обеспечивают раскручивание примерно одного витка спирали ДНК, т.е. образуется транскрипционная вилка, в которой матрица доступна для инициации синтеза цепи РНК (рис. 4-30).
После того как синтезирован олигонуклеотид из 8-10 нуклеотидных остатков, σ-субъединица отделяется от РНК-полимеразы, а вместо неё к молекуле фермента присоединяются несколько факторов элонгации.

Рис. 4-27. Строение транскриптона.
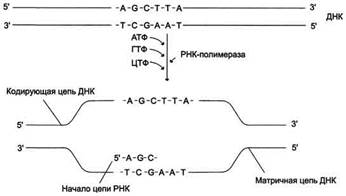
Рис. 4-28. Транскрипция РНК на матричный цепи ДНК.
Синтез РНК всегда происходит в направлении 5' → 3'.

Рис. 4-29. Строение промотора эукариотов. Промоторные элементы - специфические последовательности нуклеотидов, характерные для любого промотора, связывающего РНК-полимеразу. Первый промоторный элемент - последовательность АТАТАА- (ТАТА-бокс) отделён от сайта начала транскрипции приблизительно на 25 пар нуклеотидов (п.н.). На расстоянии примерно 40 (иногда до 120) п.н. от него располагается последовательность GGCCAATC- (СААТ-бокс).
Элонгация
Факторы элонгации повышают активность РНК-полимеразы и облегчают расхождение цепей ДНК. Синтез молекулы РНК идёт от 5'- к З'-концу комплементарно матричной цепи ДНК. На стадии элонгации, в области транскрипционной вилки, одновременно разделены примерно 18 нуклеотидных пар ДНК. Растущий конец цепи РНК образует временную гибридную спираль, около 12 пар нуклеотидных остатков, с матричной цепью ДНК. По мере продвижения РНК-полимеразы по матрице в направлении от 3'- к 5'-концу впереди неё происходит расхождение, а позади - восстановление двойной спирали ДНК.
Терминация
Раскручивание двойной спирали ДНК в области сайта терминации делает его доступным для фактора терминации. Завершается синтез РНК в строго определенных участках матрицы - терминаторах (сайты терминации). Фактор терминации облегчает отделение первичного транскрипта (пре-мРНК), комплементарного матрице, и РНК-полимеразы от матрицы. РНК-полимераза может вступить в следующий цикл транскрипции.

Рис. 4-30. Стадии транскрипции. 1 - присоединение ТАТА-фактора к промотору. Чтобы промотор был узнан РНК-полимера-зой, необходимо образование транскрипционного комплекса ТАТА-фактор/ТАТА-бокс (промотор). ТАТА-фактор остаётся связанным с ТАТА-боксом во время транскрипции, это облегчает использование промотора многими молекулами РНК-полимеразы; 2 - образование транскрипционной вилки; 3 - элонгация; 4.- терминация.
|
|
|
