 |


Классификация типов органического покоя семян
|
|
|
|
Классификация типов органического покоя семян
| Формула покоя | Тип покоя | Причины покоя | Условия нарушения покоя | Примеры |
| Типы экзогенного покоя (А) | ||||
| Собственно экзогенные: | Тормозящее действие околопл.: | |||
| a1 | слабый | слабое | Удаление или повреждение околопл., промывание плодов, длительная тепл. или хол. стратиф., некоторые другие термические воздействия | Fraxinus japonica Bliime |
| А2 | сильный | сильное | То же (кроме промывания плодов) | Pistacia vera L. |
| Аф | Физический | Водонепроницаемость покровов | Скариф., термическое воздействие, обработка H2SO4 и т. д. | Gleditsia triacanthos L., Robinia pseudoacacia L. |
| Типы эндогенного покоя (Б и В) | ||||
| Б | Морфологический | НЗ | Тепл. стратиф | Annona crassiflora Mart. |
| В | Физиологические: | ФМТ прорастания | ||
| В1 | неглубокий | ФМТ слабый | Повреждение покровов, короткая хол. стратиф., сух. хр., проращивание на свету, обработка ГБ и т. д. | Triticum aestivum L., Pinus sylvestris L., Impatiens balsamina L. |
| В2 | промежуточный | ФМТ средний | Длительная хол. стратиф. и иногда другие воздействия | Acer pseudoplatanus L. (без околопл. ), Fraxinus viridis Michx. (без околопл. ) |
| В3 | глубокий | ФМТ сильный | Только длительная хол. стратиф. (В некоторых случаях обработка ГБ или ЦТК сокращает стратиф., расширяет ее температурные границы и даже заменяет ее) | Malus domestica Borkh., Acer tataricum L., Impatiens parviflora DC. |
| Формула покоя | Тип покоя | Причины покоя | Условия нарушения покоя | Примеры | ||
| Б-В | Морфофизиологичеекие простые: | Сочетание НЗ и ФМТ прорастания | ||||
| Б-В1 | неглубокий | ФМТ слабый | Двухэтапная стратиф.: затем хол. или обработка ГКз тепл., | Actinidia kolomikta Maxim. | ||
| Б-В2 | промежуточный | ФМТ средний | То же, иногда обработка ГК3 | Aralia mandshurica Riipr. et Max. | ||
| Б-Вз | глубокий | ФМТ сильный | То же, но обработка ГКз стимулирует только доразвитие зар. | Panax ginseng C. A. Mey., Euonymus eurnpaea L. | ||
| Б—Вэ3 | Морфофизиологический глубокий эпикотильный | Сочетание НЗ и сильного ФМТ развития эпикотиля | Двухэтапная стратиф.: в тепле с. прорастают и образуются корни, затем хол. стратиф. для появления способности к росту побега | Viburnum opulus L. (без косточки) | ||
| Б —Вд3 | Морфофизиологический глубокий двойной | Сочетание НЗ и сильного ФМТ прорастания и развития эпикотиля | Четырехэтапная стратиф.: тепл. — для доразвития зар., хол. — для прорастания, опять тепл. — для роста корня, опять хол. — для появления способности к развитию эпикотиля | Trillium erectum L. | ||
| БB—В | Морфофизиологические сложные: | Сочетание НЗ и ФМТ доразвития зар. и прорастания | ||||
| БВ — В1 | неглубокий | ФМТ слабый | Хол. стратиф. или обработка ГК3 | Aralia continentalis Kitag. | ||
| БВ—В2 | промежуточный | ФМТ средний | Хол. стратиф. и в некоторой мере обработка ГКз | |||
| БВ —В3 | глубокий | ФМТ сильный | Только хол. стратиф. | Magnolia kobus DC. | ||
| Типы комбинированного покоя (А — Б — В)
| ||||||
| Различные сочетания типов экзогенного и эндогенного покоя | Сложная предпосевная подготовка или длительная стратиф. | |||||
Типы экзогенного покоя семян (А)
Прорастание семян, характеризующихся тем или иным типом экзогенного покоя, происходит в естественных условиях боль-шеи частью неодновременно и нередко растягивается на несколько лет, образуя так называемые мертвые посевы.
Следует различать прежде всего тины собственно экзогенного покоя, которые связаны со свойствами главным образом околоплодника. Долгое время считалось, что он вызывает задержку прорастания либо вследствие высокого содержания в нем ингибиторов роста, либо из-за механического препятствия росту зародыша. Соответственно М. Г. Николаева [109, 111] в своих ранних классификациях выделяла типы химического (Ах) и механического (Ам) покоя. Пока природа тормозящего влияния околоплодника мало изучена. Однако к настоящему времени накопилось довольно много наблюдений, свидетельствующих о том. что действие даже твердого околоплодника объясняется не только механическими препятствиями.
|
|
|
Предполагается, что он задерживает прорастание семян в силу комплекса причин: присутствия в нем ингибиторов, затруднения вымывания ингибиторов из семени, создания вокруг зародыша неблагоприятного осмотического давления, затруднения поступления воды в семя и сильного ухудшения газообмена зародыша. Доля участия этих факторов в задержке прорастания семян несомненно варьирует у разных видов.
В зависимости от степени тормозящего действия околоплодника на прорастание семян следует различать типы собственно экзогенного покоя слабого (А1) и сильного, (А2). У семян, находящихся в слабом эндогенном покое, задержка прорастания связана в основном с присутствием в околоплоднике ингибиторов. Они найдены в околоплоднике большого количества видов [420]. Химическая природа этих ингибиторов изучена довольна слабо. Однако ясно, что в их число входят различные фенолы и абсцизовая кислота [377, 499]. В большинстве случаев эти вещества. по-видимому, не оказывают заметного тормозящего действия на прорастание семян. Между тем у ряда видов, особенно из областей с резкой сменой дождливых и засушливых периодов года (у ясеня японского, ферулы дурнопахнущей), ингибиторы околоплодника предотвращают прорастание семян в неблагоприятные для растений периоды года. Как правило, для того чтобы устранить такую задержку прорастания, достаточно адсорбции ингибирующих веществ почвой в естественных условиях, искусственного удаления околоплодника или обильного промывания плодов. В торможении прорастания семян, характеризующихся слабым экзогенным покоем, известную роль играют и другие свойства околоплодника, перечисленные выше. В отличие от сильного экзогенного покоя при слабом покое тормозящее действие околоплодника проявляется не всегда.
Сильный экзогенный покой (А2) чаще всего наблюдается У семян, заключенных в твердую косточку, которая сильно задерживает прорастание многих плодовых, орехоплодных и других растений. Наблюдения на сливе (Primus), боярышниках (Сrаtaegus), фисташке (Pistacia vera), калине (Viburnum opulus) и некоторых других растениях, приводят к выводу, что твердая косточка, представляющая собой внутреннюю часть околоплодника — эндокарний, не только или, вернее, не столько является механическим препятствием прорастанию, сколько оказывает на этот процесс тормозящее действие по причинам, перечисленным выше. Действительно, установлено, что у некоторых видов в косточке содержатся ингибиторы, высказаны предположения, что она может препятствовать вымыванию ингибиторов из семян или создавать вокруг них неблагоприятные осмотические условия. Определения дыхания показали, что у миндаля и фисташки косточка, даже открытая, сильно затрудняет газообмен семян.
|
|
|
Удаление околоплодника производится в небольших партиях семян вручную. В больших партиях рекомендуются различные методы обработки косточек (см. гл. 2). Нередко тормозящее действие околоплодника преодолевается длительной стратификацией, способствующей его разрушению.
Особенно сильно затруднено прорастание семян, находящихся в физическом покое (Аф), который нередко обозначается также термином «твердосемянность» и объясняется полной водонепроницаемостью кожуры, реже — околоплодника. Физический покой характерен для семян многих хозяйственно важных растений, поэтому на нем следует остановиться несколько подробнее.
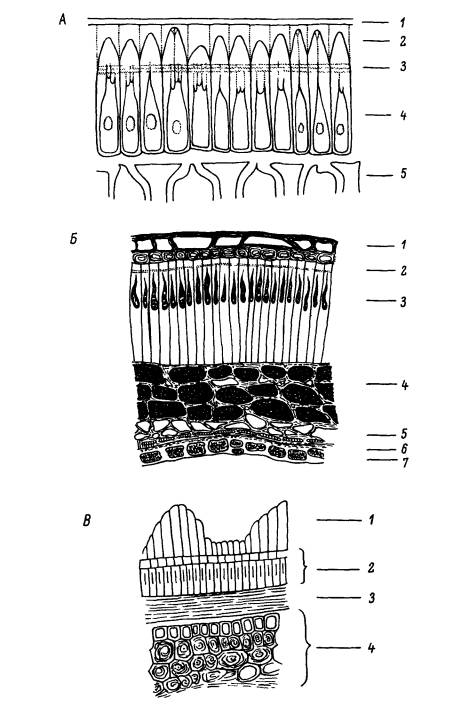
Явление твердосемянности широко распространено в растительном мире и свойственно многим представителям ряда семейств: мимозовых (Mimosaceae), цезальпиниевых (Caesalpiniaceae), мотыльковых (Fabaceae), мальвовых (Malvaceae), липовых (Тiliасеае), стеркулиевых (Sterculiaceae), канновых (Саппасеае) и некоторых других. А. В. Попцов [142] рассматривает твердосемянность как особый тип покоя, принципиальное отличие которого от остальных типов состоит в том, что в этом случае имеют место не частичное затруднение поступления воды и подавление ростовых процессов, а полная их остановка. Физический покой вызывается особенностями строения семенной кожуры (рис. 1): наличием кутикулы и сильно развитой палисадной эпидермы — слоя вытянутых клеток, который пересекают одна или несколько световых линий.
|
|
|
А — Melilotus albus Medic.: 1 — кутикула, 2 — колпачки палисадных клеток, 3 — световые линии, 4 — палисадный слой, 5 — гиподерма; Б — Abutilon avicennae Gaertn.: 1 — производное внешнего слоя интегумента, 2 — световые линии, 3 — палисадный слой, 4 — пигментный слой, 5 — паренхимные клетки, 6 — бахромчатые клетки, 7 — алейроновый слой: В — Convolvulus arvensis L.: 1 — эпидермис, 2 — палисадный слой, 3 — внутренняя кутикула, 4 — эндосперм.
Состояние твердосемянности развивается постепенно, по мере высыхания семян на последних фазах созревания или во время хранения после отделения их от материнского растения. Вначале семена отдают влагу всей поверхностью кожуры, но с развитием твердосемянности потеря влаги происходит через рубчик. Последний обладает очень важной особенностью: он остается открытым в сухом воздухе, но при повышенной влажности атмосферы моментально закрывается. Таким образом, твердые семена постепенно теряют воду, как и семена с поврежденной кожурой, но в отличие от последних лишены возможности поглощать из воздуха парообразную влагу ( рис. 2 ).

Процент твердых семян сильно варьирует в зависимости от видовых особенностей, а также условий созревания и хранения. В частности, низкая влажность воздуха ведет к резкому увеличению процента твердых семян. А. В. Попцов приводит следующие примеры: у видов кассии (Cassia) в Средней Азии были твердыми 10—30 % семян, а в Батуми всего 2—3 %. Столь же резко различались семена алтея (Althaea officinalis) из Дагестана и Украины.
Семена, в которые не проникает вода, естественно не могут прорастать. Когда твердосемянность нарушается, семена приобретают способность к набуханию и при благоприятных условиях быстро прорастают, так как их зародыши обычно HP находятся в покое. В естественных влажных условиях у некоторых видов семена постепенно и очень медленно утрачивают твердость; этот процесс может продолжаться много лет. Наряду с этим быстрое прорастание твердых семян нередко провоцируют различные температурные и биотические воздействия.
Основным фактором, выводящим твердые семена из состояния физического покоя, являются температурные воздействия, при этом положительное влияние оказывают как низкие, так и высокие и переменные температуры. Сильное влияние на увеличение способности к набуханию семян травянистых бобовых оказывает зимнее понижение температуры. Эффект усиливается, если имеют место сильные колебания температуры, и особенно при повышенной влажности воздуха. У семян канатника (Abutilon) процент набухания был тем выше, чем дольше длилось промораживание и чем ниже была температура (в испытанных пределах от -11 до -23°). На семена бобовых слабые морозы (от -5 до -8°) не оказывали действия, между тем понижение температуры до -10° и ниже приводило к значительному увеличению процента набухающих семян. В то же время периодическое промораживание семян повилики (Cuscuta) не дало эффекта.
|
|
|
Немаловажное влияние оказывает на твердые семена повышение температуры как во время хранения, так и при проращивании. Прогревание семян канатника в течение 6—8 ч при 40° повысило их всхожесть до 89—95 %. Прогревание семян, особенно при меняющихся обычно в течение суток температурах, приводит, как видно из табл. 2, к утрате твердости и у различных бобовых. В этом опыте изменение температуры в течение дня производилось постепенно, как это происходит в естественных условиях в летний день на почве. Большое значение имеет температура проращивания. При высоких и переменных температурах твердые семена ряда видов бобовых набухают более успешно, чем на холоде.
|
|
|
