 |


Структурная организация фотосинтетического аппарата, строение листа как органа фотосинтеза, ультраструктура и онтогенез хлоропластов.
|
|
|
|
Основой фотосинтетического аппарата служат:
− у высших зел еных растений внутриклеточные органеллы – хлоропласты;
− у большинства водорослей − специализированные внутриклеточные органеллы – хроматофоры;
− у фотосинтезирующих бактерий и синезел еных водорослей – тилакоиды (мембраны их содержат пигмент бактериохлорофилл или бактериовиридин, а также др. компоненты реакций фотосинтеза), погруженные в периферические слои цитоплазмы.
Хлоропласты занимают 20–30% объ ема растительной клетки. У водорослей, например хламидомонады, имеется один хлоропласт, в клетке высших растений содержится от 10 до 70 хлоропластов и более.
Хлорофилл и каротиноиды, погруженные в тилакоидные мембраны хлоропластов, собраны в функциональные единицы – фотосистемы, каждая из которых содержит примерно 250 молекул пигментов. Фотосистема – совокупность ССК (светособирающего комплекса), фотохимического реакционного центра и переносчиков электрона.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикрепл енные к ним фикобилисомы – палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины, за ними фикоцианины и аллофикоцианины, последовательно передающие энергию на хлорофилл a реакционного центра.
Роль ССК состоит в том, чтобы собирать и передавать энергию квантов света на небольшое количество молекул (молекулы-ловушки) реакционных центров П680 и П700 которые и осуществляют фотохимические реакции.
Органом фотосинтеза у высших растений является лист. Кроме фотосинтеза в жизни растений лист выполняет функции газообмена, транспирации, синтеза ряда органических веществ, в том числе и фитогормонов.
|
|
|
Листья, будучи боковыми органами, как правило, имеют более или менее плоскую форму, что способствует созданию максимальной фотосинтезирующей поверхности.
Основной тканью листа является паренхимный комплекс клеток, несущий хлоропласты – мезофилл (греч.mesos – средний и филл) (рисунок 1). Остальные ткани листа обеспечивают работу мезофилла и поддерживают связь с окружающей средой. Покровная ткань (эпидермис) регулирует газообмен и транспирацию, защищает лист от внешних воздействий. Проводящие ткани осуществляют отток и приток веществ, поддерживают нормальное оводнение фотосинтезирующих клеток. Механические ткани совместно с живыми тургесцентными клетками мезофилла и эпидермиса образуют опорную систему листа.
Мезофилл. Представлен клетками основной паренхимы, расположенными между верхним и нижним эпидермисом (исключая механические ткани и проводящие пучки). Состоит из живых клеток, с тонкими оболочками, округлой или слегка вытянутой формы, реже с небольшими выростами. Благодаря многочисленным хлоропластам мезофилл окрашен в зеленый цвет, так называемая хлоренхима листа. Иногда оболочки клетки образуют складки внутрь полости клетки, что увеличивает рабочую поверхность хлоренхимных клеток.
У типичных дорсовентральных листьев мезофилл неоднороден и диффе-ренцирован на столбчатую (палисадную) и губчатую паренхиму.
Клетки столбчатой паренхимы плотно сомкнуты, имеют продолговатую форму и располагаются перпендикулярно к верхней стороне листа. Столбчатый мезофилл может быть однослойный, иногда двуслойный, редко (у светолюбивых растений) многослойный. Основная функция палисадной паренхимы – ассимиляция углекислого газа (фотосинтез). Палисадная ткань располагается в наилучших условиях освещения, непосредственно под верхней эпидермой. Благодаря тому, что ее клетки вытянуты перпендикулярно к поверхности листа, у них имеется возможность регулировать направление и расположение хлоропластов таким образом, чтобы избежать повреждающего действия прямой солнечной радиации на фотосинтезирующий аппарат.
|
|
|
Губчатую паренхиму составляют относительно округлые клетки с большими межклетниками, которые занимают больший объем, чем сами клетки. Рыхлая структура мезофилла обусловливает большую общую площадь поверхности клеток, обращенной к межклетникам. Через крупные межклетники губчатого мезофилла идет газообмен.
Хлоропласты. Форма – овальные тельца длиной 5-10 мкм и диаметром 2-3 мкм. Количество в клетке листа – 15-20, у некоторых водорослей - 1-2 гигантских хлоропласта различной формы. Окраска – зеленая, обусловлена содержанием в хлоропластах хлорофилла.
Хлоропласты – двухмембранные органоиды (рисунок). Внутреннее содержимое хлоропластов − строма (матрикс) представляет собой гомогенную среду. Хлоропласты, как и другие органоиды, например, митохондрии, имеют ламеллярное строение, т.е. внутренние структуры органоида образованы инвагинациями внутренней мембраны – ламеллами. В хлоропластах внутренняя мембрана образует уплощенные инвагинации — тилакоиды, которые могут иметь форму дисков и в этом случае называются тилакоидами гран.
Несколько лежащих друг над другом тилакоидов образуют стопку – грану. Другие тилакоиды, связывающие между собой граны или не контактирующие с ними, называются тилакоидами стромы. При таком строении значительно увеличивается фотоактивная поверхность хлоропластов и обеспечивается максимальное использование световой энергии. В мембранах тилакоидов локализованы зеленые (хлорофиллы), желтые и красные (каротиноиды) пигменты, компоненты редокс-цепей и запасания энергии, участвующие в поглощении и использовании энергии света.
Граны часто имеют форму цилиндров размером 2 мкм, в которых пигментно-липидные слои чередуются с белковыми слоями. Морфологически тилакоиды не гомогенны. На внутренней поверхности их мембран имеются специфические структурные образования, названные Парком квантосомами.
Хлоропласты – системы, способные к автономному синтезу белков. В них присутствуют низко- и высокомолекулярная РНК, специфическая кольцевая ДНК и ферменты, активирующие аминокислоты. Хлоропласты обладают собственными рибосомами.
|
|
|
Функции хлоропластов:
1) осуществление фотосинтеза (основная функция);
2) в хлоропластах происходит вся сложная цепь процессов превращений первичных продуктов фотосинтеза (наращивание углеродной цепи, образование и распад полимерных форм углеводов и др.);
Биохимические системы синтеза и превращения углеводов функционируют в строме хлоропластов. В ней же может откладываться крахмал. В хлоропластах представлен весь набор биохимических систем, участвующих в синтезе АТФ.
Онтогенез хлоропластов. При нормальном освещении пропластиды превращаются в хлоропласты. Сначала они растут, при этом происходит образование продольно расположенных мембранных складок от внутренней мембраны. Одни из них простираются по всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых хлоропластов.
Несколько иначе развитие пластид происходит в темноте. У этиолированных проростков происходит в начале увеличение объема пластид – этиопластов, но система внутренних мембран не строит ламеллярные структуры, а образует массу мелких пузырьков, которые скапливаютсяя в отдельные зоны и даже могут формировать сложные решетчатые структуры (проламеллярные тела). В мембранах этиопластов содержится протохлорофилл, предшественник хлорофилла желтого цвета. Под действие света из этиопластов образуются хлоропласты, протохлорофилл превращается в хлорофилл, происходит синтез новых мембран, фотосинтетических ферментов и компонентов цепи переноса электронов.
27. Дыхание и обмен веществ в растительной клетке. Взаимосвязь фотосинтеза и дыхания. Использование промежуточных продуктов окисления дыхательного субстрата в процессах биосинтеза
Тесная связь дыхания с биосинтетическими функциями клетки не ограничивается использованием реализуемой в ходе окислительных процессов химической энергии дыхательного субстрата. Промежуточные продукты дыхания могут использоваться в процессах новообразования компонентов протоплазмы. Например, аминирование α-кетоглутаровой кислоты приводит к образованию глутаминовой кислоты.
|
|
|

К аминированию путем непосредственного присоединения аммиака способны также ПВК и ЩУК. Так, прямое аминирование ПВК приводит к образованию аланина.
В процессах восстановительного биосинтеза используется почти исключительно НАДФ-Н, тогда как НАД-Н потребляется преимущественно для генерирования АТФ.
С окислительными превращениями трикарбоновых кислот в цикле Кребса тесно связаны процессы биосинтеза жиров. Конденсация двух молекул уксусной кислоты при участии КоА приводит к образованию ацетоуксусной кислоты, при восстановлении которой получается масляная кислота.
Путем последовательного присоединения ацетильных радикалов, активируемых КоА, и происходит наращивание углеродной цепи - синтез жирных кислот в организме.
В ходе гексозомонофосфатного дыхания образуются активные формы пентоз, участвующие в построении важнейших соединений клетки. У гетеротрофных организмов гексозомонофосфатное Дыхание представляет единственный путь в образовании пентоз.
Таким образом, промежуточные продукты дыхания могут дать начало всем основным компонентам растительной клетки, и дыхание в этом случае выполняет функции, которые по своему значению для синтетической функции клетки не уступают фотосинтезу.
Связь дыхания и фотосинтеза. Фотосинтез и дыхание тесно связаны. Например, если рассматривать на молекулярном уровне одинаковые начальные и конечные этапы цикла Кальвина и пентозофосфатного пути, многие промежуточные продукты и каталитические системы тождественны. Промежуточные продукты обоих процессов широко используются в различных биосинтезах. Основными функциями как митохондрий, так и хлоропластов являются транспорт электронов и сопряженные с переносом электронов процессы связывания энергии в видеАТФ при фосфорилировании.
При фотосинтезе активирование электрона происходит за счет энергии кванта света, при дыхании - за счет энергии химических связей, сосредоточенной в молекуле дыхательного субстрата. Однако свободная энергия любого органического соединения генетически связана с фотосинтезом. Следовательно, как при фотосинтезе, так и при дыхании источник образования активного электрона - это электромагнитная энергия кванта света.
Много общего обнаруживают компоненты ЭТЦ митохондрий и хлоропластов. Последнее позволяет считать, что при формировании каталитических систем аэробного дыхания, возникших в эволюционном отношении вслед за фотосинтезом, были широко использованы важнейшие компоненты ЭТЦ фотосинтеза.
|
|
|
Ультраструктура и химический состав органелл не только принципиально близки, но и во многих отношениях общие. Обе органеллы способны самостоятельно осуществлять синтез белков. Это свидетельствует о большой автономности этих органелл и способности к самовоспроизведению, подтверждает идею об их происхождении от общего предшественника - пропластиды. Кроме того, практически все продукты процесса гликолиза, цикла Кребса и гексозомонофосфатного пути обычно обнаруживаются в группе соединений, относящихся к самым ранним продуктам фотосинтеза.
Скорость как дыхания, так и фотосинтеза может служить показателем функциональной стойкости растений, и их измерение позволяет выявить причины изменений продуктивности в конкретных условиях среды.
Гуттация, транспирация (устьичная и кутикулярная); физиологическое значение этих процессов, количественные показатели транспирации (интенсивность и продуктивность транспирации, транспирационный коэффициент).
Гуттация наблюдается при высокой влажности воздуха. В результате деятельности нижнего концевого двигателя на концах и зубчиках листьев выделяется капельно-жидкая влага − растение гуттирует. Это явление характерно для листьев тропических растений, приспособленных к жизни в условиях повышенной влажности, при которых транспирация затруднена.
Транспира́ция — это испарение воды растением. Основным органом транспирации является лист. Вода испаряется с поверхности листьев через клеточные стенки эпидермальных клеток и покровные слои (кутикулярная транспирация) и через устьица(устьичная транспирация).
В результате потери воды в ходе транспирации в клетках листьев возрастает сосущая сила. Это приводит к усилению поглощения клетками листа воды из сосудов ксилемы и передвижению воды по ксилеме из корней в листья. Таким образом, верхний концевой двигатель, участвующий в транспорте воды вверх по растению, обусловлен транспирацией листьев.
Кутикулярная транспирация[править
Снаружи листья имеют однослойный эпидермис, внешние стенки клеток которого покрыты кутикулой и воском, образующие эффективный барьер на пути движения воды. На поверхности листьев часто развиты волоски, которые также влияют на водный режим листа, так как снижают скорость движения воздуха над его поверхностью и рассеивают свет и тем самым уменьшают потери воды за счет транспирации.
Интенсивность кутикулярной транспирации варьирует у разных видов растений.
Устьичная транспирация[править | править вики-текст]
Устьица представляют собой щель в подъустьичную полость, окаймленную двумя замыкающими клетками серповидной формы. Устьица играют важную роль в газообмене между листом и атмосферой, так как являются основным путем для водяного пара, углекислого газа и кислорода. Устьица находятся на обеих сторонах листа. В среднем число устьиц колеблется от 50 до 500 на 1 мм². Транспирация через устьица идет почти с такой же скоростью, как и с поверхности чистой воды. Это объясняется законом И. Стефана: через малые отверстия скорость диффузии газов пропорциональна не площади отверстия, а диаметру или длине окружности.
Транспирация играет в жизни растения значительную роль:
1. Она служит одним из основных механизмом передвижения воды и различных веществ вверх по стеблю.
2. оказывает большое влияние на процессы воздушного питания растения.
3. Защищает листья от перегрева.
4. Снижение интенсивности транспирации в результате недостатка воды существенно сдвигает обычный ход физиологических процессов в листе. Оно сопровождается чрезмерным повышением температуры листьев, нарушениями в коллоидной системе протоплазмы, подавлением фотосинтеза, усилением дыхания и т.д.
Интенсивность транспирации — это количество воды, испаряемой растением в граммах за единицу времени в часах единицей поверхности в дм².
Продуктивность транспирации -это количество граммов сухого вещества, накопленного в растении при испарении 1000 г воды.
Транспирационный коэффициент количество воды (в граммах), расходуемое на образование 1 г сухого вещества. Т. к. зависит от климатических и почвенных условий и от вида растений (например, у просовидных злаков он относительно низок
29. Фитогормоны и их функции. Ауксины, гиббереллины и цитокинины – основные горомны роста. Абсцизовая кислота и этилен – их функции в растенях. Использование фитогормонов и других физиологически активных веществ в растениеводстве.
Фитогормоны – низкомолекулярные органические вещества, вырабатываемые растениями и имеющие регуляторные функции.
Фитогормоны имеют следующие общие черты:
§ действие проявляют в очень низких концентрациях (10-13- 10-5 моль/л);
§ как правило, образуются в одной части растения, а действуют в другой, куда транспортируются;
§ образуются в отдельных частях растения, но распространяются по всему организму, составляя своеобразное гормональное поле;
§ регулируют крупные морфогенетические и физиологические программы и подпрограммы.
Основные фитогормоны: ауксин, гиббереллины, цитокинины, абсцизовая кислота и этилен.
Ауксин (ИУК) синтезируется растущими апикальными зонами стеблей, в том числе молодыми листьями. От апекса ауксин мигрирует в зону растяжения, где он специфически влияет на рост растяжением. Транспорт ауксина происходит полярно со скоростью 10-15 см/ч от вершины побегов к корням.

Гиббереллины – фитогормоны, преимущественно класса тетрацикличес-ких дитерпеноидов. Гиббереллины синтезируются особенно интенсивно в растущих апикальных стеблевых почках растений, в хлоропластах листьев, в формирующихся семенах, в зародыше прорастающих семян. Гиббереллины передвигаются в растениях неполярно, пассивно вверх-вниз и латерально.
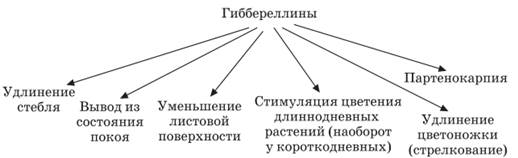
Цитокинины. Основное место синтеза - в апикальной меристеме корней. Образуются они также в молодых листьях и почках, в развивающихся плодах и семенах. Цитокинины обнаружены в галловых опухолях на листьях растений, возникших под действием паразитических грибов и насекомых. Цитокинины транспортируются с водным током по сосудам ксилемы из апексов корней в надземные части растения со скоростью до 10-50 см/ч.

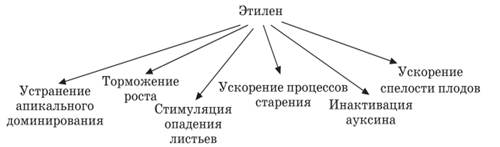
Этилен (СН2 = СН2) – гормон старения (гормональный фактор газоподобного типа).
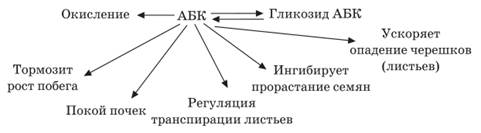
Абсцизовая кислота (АБК) – природный гормональный ингибитор роста терпеноидной природы. АБК синтезируется главным образом в листьях, а также в корневом чехлике. Перемещение АБК в растениях происходит как в базипетальном, так и в акропетальном направлениях в составе ксилемного и флоэмного соков. Этот гормон чаще выступает как антагонист ИУК, цитокинина и гиббереллинов.
Использование фитогормонов и других физиологически активных веществ в растениеводстве.. Фитогормоны и синтетические регуляторы роста широко используют в сельском хозяйстве. Некоторые из применяющихся веществ служат синтетическими аналогами природных регуляторов. Созданы и многие другие синтетические регуляторы роста, не имеющие природных аналогов, меняющие гормональный статус растений и как результат изменяющие течение физиологических процессов в желательном для земледельца направлении.
|
|
|
