 |


Фибрилляции желудочков п эктопическую активность сердца при
|
|
|
|
постинфарктном кардиосклерозе (M ± m)
| Вариант опытов | Исходная ЧСС, ударов в минуту | Пороговое напряжение тока, В | — Δ ЧСС, ударов в минуту | Суммарное число экстрасистол | Порог фибрилляции желудочков, мА | |||
| 1 порог | 2 порога | 3 порога | 4 порога | |||||
| Контроль (11) | 389+11 | 0,4 + 0,06 | 40+3,6 | 122 + 21 | 170+17 | 193±16 | 6,8+0,7 | |
| Вальпроат | 379±8 | 0,3 + 0,05 | 55±7 | 196 + 29 | 208±25 | 223+25 | — | 6,4±0,7 |
| натрия (10) Постинфарктный кардиосклероз (11) | 380+13 | 0,3 + 0,03 | 44±5 | 145+16 | 176±18 | 205+17 | 2,3±0,3* | |
| Постинфарктный кардиосклероз + вальпроат натрия (10) | 390±12 | 0,3±0,03 | 44±4 | 143±18 | 195±24 | 207±24 | 5,3+0,3** | |
| Примечание. Цифры в скобках — число животных. * Отличия от варианта «контроль» достоверны. ** Отличия от варианта «постинфарктный кардиосклероз» достоверны. |
ческого порога фибрилляции сердца и его эктопической активно^ сти изучали описанными выше методами, используя ту же дозу вальпроата натрия, что и при острой ишемии. Установили, что вальпроат натрия увеличивает отрицательно хронотропный эффект блуждающего нерва, но при этом на фоне увеличенной по степени брадикардии отсутствуют одиночные экстрасистолы, которые наблюдались у животных, не получавших препарата.
У животных с кардиосклерозом, как и в ранее описанных экспериментах, наблюдалось значительное — почти в 3 раза снижение порога фибрилляции сердца, а на фоне вагусной брадикардии возникали многочисленные экстрасистолы — 401 на группу обследованных животных. Вальпроат натрия в 2,5 раза повысил порог фибрилляции и в 10 раз уменьшил число экстрасистол. Таким образом, этот «накопитель ГАМК» почти полностью устранил нарушения электрической стабильности сердца, типичные для постинфарктного кардиосклероза. У интактных животных не отмечалось какого-либо влияния препарата на сердечный ритм или порог фибрилляции, явления ишемии и сопутствующего стресса при достаточно «старом» постинфарктном кардиосклерозе отсутствовали. Поэтому возникает подозрение, что при таком кардиосклерозе рубцу в сердечной мышце соответствует очаг повышенной возбудимости в центрах головного мозга, регулирующих функцию сердца. Этот очаг за счет усиления адренергических влияний на сердце или иным путем приводит к обнаруженным нарушениям электрической стабильности. Подавление очага избытком ГАМК устраняет эти нарушения по существу на основе того же механизма, что и эпилептические приступы. Разумеется, это предположение требует дальнейшей проверки. Однако оно согласуется с прямыми данными Г. Н. Крыжановского и Ю. И. Пивоварова (1982) о том, что в результате блокады ГАМК-рецепторов пенициллином в определенных структурах головного мозга может происходить формирование очагов возбуждения, которые сочетаются с возникновением сердечных аритмий. Аритмии исчезают вместе с исчезновением стойких очагов возбуждения.
|
|
|
В целом, полученные данные свидетельствуют, что вальпроат натрия антиконвульсивное средство, до последнего времени не использовавшееся в кардиологии, обладает выраженной способностью предотвращать аритмии, фибрилляцию и остановку сердца, а также устранять нарушения электрической стабильности сердца при постинфарктном кардиосклерозе.
Этот факт согласуется с представлением о важной роли ГАМК-ергической стресс-лимитирующей системы в предупреждении фибрилляции и остановки сердца при стрессе, ишемии и кардиосклерозе. Врожденная или приобретенная неполноценность этой системы, по-видимому, может играть важную роль в патогенезе аритмий и острой сердечной смерти. Повышение активности ГАМК-ергической системы, напротив, является вероятным фактором, определяющим защитный, антиаритмический эффект адаптации к стрессорным ситуациям окружающей среды.
|
|
|
|
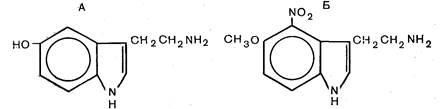
| Рис. 23. Структурные формулы серотонина (А) и его аналога П-109 (Б). Пояснение в тексте. |
Другой центральной стресс-лимитирующей системой, которая может играть роль в защитном эффекте адаптации при аритмии и фибрилляции сердца, вызванных острой ишемией, является серотонинергическая система головного мозга. Действительно, в настоящее время имеется ряд фактов, указывающих на роль серотонинергической системы в этом защитном эффекте. Прежде всего наши опыты показали, что рассмотренная выше адаптация к повторным стрессорным воздействиям приводит к увеличению содержания серотонина в структурах головного мозга. Так показано, что в среднем мозге, где в области шва расположены основные серотонинергические нейроны, содержание серотонина, определявшееся известным флуорометрическим методом [Maickel R., Miller E., 1966], возросло с 1061 нг/г до 1696 нг/г, т. е. в среднем на 60% (р<0,05). Кроме того, ранее установлено, что такое же накопление серотонина в мозге, вызванное с помощью сложного комплекса фармакологических средств, предотвращает снижение порога фибрилляции сердца при острой ишемии [Lown В. et al., 1980]. На этом основании мы решили использовать синтетические аналоги серотонина, не применявшиеся ранее в кардиологии, для предупреждения фибрилляции сердца при острой ишемии. В качестве такого аналога был избран хлоргидрат 4-нитро-5-метокситриптамина (П-109), синтезированный И. А. Петруниным в Московском химико-технологическом институте им. Д. И. Менделеева. Это вещество в основе своей так же, как и серотонин, имеет индольное кольцо, но отличается от серотонина наличием метоксигруппы в 5-м положении и нитрогруппы — в 4-м положении; оно обладает радиопротекторным действием и низкой токсичностью. На рис. 23 приведена структурная формула П-109. Антиаритмический эффект этого аналога серотонина оценивали на модели острой ишемии, воспроизводившейся на бодрствующих животных при закрытой грудной клетке по методу L. Lepran и соавт. (1983). Использованный аналог вводили за 30 мин до коронароокклюзии в дозе 50 мг/кг внутрибрюшинно. Результаты исследований показали, что предварительное введение П-109 в 4 раза снижало частоту фибрилляции сердца и смертность животных во время 10-минутной коронароокклюзии и более чем в 20 раз уменьшало суммарную длительность аритмий. Таким образом,
|
|
|
это вещество обладает выраженным антиаритмическим действием при острой ишемии и может быть рекомендовано для клинических испытаний.
В целом, представленные выше результаты свидетельствуют, что, избирательно активируя центральные стресс-лимитирующие системы или вводя синтетические аналоги метаболитов этих систем, можно защитить организм от повреждений, в патогенезе которых важную роль играет стресс-реакция, и в частности, от фибрилляции сердца.
Свободнорадикальное окисление в патогенезе ишемических и стрессорных повреждений миокарда и кардиопротекторное действие антиоксидантов 1
В ходе естественного процесса адаптации к коротким стрессорным воздействиям закономерно возникает активация антиоксидантных систем организма. Данные Т.Г.Сазонтовой (1986), представленные в табл. 29, свидетельствуют, что однократный дли-
Таблица 29. Активность антиоксидантных ферментов в сердечной мышце крыс (M±m) при стрессе
| Вариант опытов | Активность ферментов | ||
| каталаза, мМ Н202/г/мин | супероксиддисмутаза, усл. ед. /г/мин | глутатионпероксидаза, мМ НАДФ. Н г/мин | |
| Контроль (4) | 1,82±0,11 | 58,7±2,1 | 19,2+0,06 |
| Стресс (4) | 1,47±0,18* | 56,1±2,1 | 18,7±0,04 |
| Адаптация (4) | 2,52±0,27* | 67,9+6,5* | 19,0±0,07 |
| Адаптация + стресс (4) | 1,96+0,20 | 64,0±2,5 | 19,4+0,03 |
| Примечание. Цифры в скобках — число животных. * Отличия от варианта «контроль» достоверны. |
тельный эмоционально-болевой стресс, который, как известно, активирует свободнорадикальное окисление, приводит к некоторому снижению активности каталазы и слабой тенденции к снижению активности других антиоксидантных ферментов в сердечной мышце; адаптация к коротким эмоционально-болевым стрессорным воздействиям, напротив, приводит к увеличению активности каталазы на 38%, супероксиддисмутазы на 16% и существенно не влияет на активность глутатионпероксидазы. Соответственно тяжелый стресс у адаптированных животных не приводит к снижению активности антиоксидантных ферментов в сердечной мышце.
|
|
|
Этот результат согласуется с ранее приведенными данными о том, что устойчивость изолированного сокращающегося пред-
1 Раздел написан совместно с В. В. Диденко.
сердия к аритмогенному действию индуктора свободнорадикального окисления липидов Н202 у адаптированных к стрессорным воздействиям и физическим нагрузкам животных существенно повышена. Они позволяют в новом свете рассматривать кардиопротекторную роль естественных антиоксидантных систем и вводимых извне антиоксидантов. Поскольку антиоксидантные системы или синтетические антиоксиданты блокируют активацию ПОЛ и защищают сердце при стрессе и ишемии [Меерсон Ф. 3., 1984], то они, очевидно, должны предупреждать или ограничивать аритмии при таких состояниях. В этом плане в дальнейшем изложении мы рассмотрим новые данные о роли ПОЛ в патогенезе стрессорных и ишемических повреждений сердца и о кардиопротекторном действии антиоксидантов.
Первые прямые данные о роли ПОЛ в патогенезе инфаркта миокарда были получены А. X. Коганом, А. М. Кудриным, С. М. Николаевым (1976), показавшими увеличение хемилюминесценции, обусловленное активацией свободнорадикальных процессов в зоне ишемии при инфаркте миокарда. Вскоре была показана роль ПОЛ в патогенезе стрессорных некоронарогенных повреждений миокарда [Меерсон Ф. 3., Каган В. Е., Прилипко Л. Л. в др., 1979].
В последние годы была выявлена роль ПОЛ в возникновении аритмий [Меерсон Ф. 3., Салтыкова В. А. и др., 1984; Woodward В., Zakaria M. N., 1985], в механизме кислородного парадокса [Guarnieri G. et al., 1980], в повреждении сердца при анемиях [Меерсон Ф. 3., Евсевьева М. Е., 1984]. Далее, появились многочисленные работы о кардиопротекторном действии естественных и синтетических антиоксидантов, предварительное введение которых, как оказалось, обладает способностью предупреждать повреждения при ишемии и стрессе и способно ограничивать зону ишемического повреждения при инфаркте миокарда [Меерсон Ф. 3., 1980; 1984; Jolly S. et al., 1984].
В итоге этих исследований роль свободнорадикального окисления в патогенезе заболеваний сердца и возможность использования антиоксидантов как кардиопротекторов привлекли широкое внимание кардиологов и по существу стали одним из важных направлений в современной кардиологии.
Детальному рассмотрению данной проблемы посвящен ряд; обзоров, суммирующих представления о роли свободных радикалов в патологии сердца и защитном действии антиоксидантов. В соответствии с имеющимися данными процессы свободнорадикального окисления постоянно протекают в миокарде. Частным проявлением свободнорадикальных процессов является ПОЛ, развивающееся в фосфолипидах мембранных структур кардиомиоцитов. Отсутствие сколько-нибудь значительного повреждения мембранных структур при этом определяется тем фактом, что клетки, обладают мощной антиоксидантной системой, которая слагается из комплекса антиоксидантных ферментов и химических антиоксидантов, т. е. витаминов Е, С, К и других соединений со свободной SH-группой (глутатион, цистеин и др.), и, наконец, дополняется так называемым структурным антиоксидантом, т. е. определенным образом детерминированной организацией липидов в биомембранах [Владимиров Ю. А., Арчаков А. И., 1972].
|
|
|
Таким образом, в основе поддержания свободнорадикального гомеостаза клеток лежит баланс между образованием и элиминацией свободных радикалов. Существенно, что устойчивость такого равновесия имеет свои границы и определяется, с одной стороны, мощностью систем антиоксидантной защиты, а с другой — интенсивностью процессов генерации радикалов. Поэтому в основе активации ПОЛ всегда лежит один из трех общих механизмов: во-первых, первичная чрезмерная генерация активных форм кислорода, превышающая физиологические возможности антиоксидантных систем клетки; такая ситуация имеет место при гипербарической оксигенации, перенасыщающей мембранные структуры кислородом и, по-видимому, объясняет также кардиотоксическое действие катехоламинов при стрессе. Во-вторых, первичное снижение мощности антиоксидантных защитных систем, наступающее, например, при авитаминозе Е, при отравлениях ферментными ядами и некоторых наследственных дефектах (акаталазия, желтуха новорожденных и др.). В-третьих, сочетание этих возможностей в случае ишемии, определяемое, с одной стороны, массивной потерей антиоксидантов, выходящих через поврежденную наружную мембрану клеток, а с другой — активной генерацией инициаторов ПОЛ.
Независимо от первичной причины, вызвавшей нарушение баланса и активацию ПОЛ, в сердечной мышце развиваются если не однотипные, то достаточно сходные повреждения клеточных структур и функции.
На основе этих общих соображений мы рассмотрим роль ПОЛ в патогенезе повреждений сердца при стрессе, ишемии, в возникновении аритмий при этих состояниях, а также возможность использования антиоксидантов для предупреждения вызываемых ПОЛ нарушений.
Стресс-реакция, как показано выше, составляет необходимое звено адаптации организма к различным факторам окружающей среды, а адренергическая мобилизация функции и метаболизма сердца — необходимый компонент этой реакции.
Однако при увеличении длительности и интенсивности стрессорных воздействий или при введении больших доз катехоламинов адаптивный эффект закономерно превращается в повреждающий. Так, после длительного — 6-часового — эмоционально-болевого стресса в сердечной мышце развивается значительная активация ПОЛ, проявляющаяся увеличением содержания гидроперекисей липидов и шиффовых оснований в миокарде в 2—3 раза [Меерсон Ф. 3., Каган В. Е., Архипенко Ю. В. и др., 1981]. Это сопровождается лабилизацией лизосом сердечной мышцы и ускоренным выходом в перфузат креатинфосфокиназы (КФК), лактатдегидрогеназы и других ферментов из миокарда изолированных сердец перенесших стресс животных [Меерсон Ф. 3., Каган В. Е., Голубева Л. Ю. и др., 1979]. Одновременно, при исследовании миокарда определяется падение активности и ускорение термоинактивации Na, К-АТФазы сарколеммы [Меерсон Ф. 3.,
Сазонтова Т. Г. и др., 1986], а также снижение активности Са-АТФазы и способности связывать Са2+ в мембранах СПР [Каган В. Е., Архипенко Ю. В. и др., 1983]. Этот первичный комплекс сдвигов надо рассматривать как результат стрессорного повреждения мембран вследствие чрезмерной активации ПОЛ, фосфолипаз и детергентного действия избытка жирных кислот и лизофосфатидов. Для оценки роли ПОЛ в этом процессе существенно, что повреждения сарколеммальной мембраны и инактивация Na, К-АТФазы, а также упомянутые повреждения саркоплазматической мембраны и инактивация Са-АТФазы закономерно воспроизводятся посредством индукции ПОЛ in vitro. Причем, изучение динамики данного процесса [Архипенко Ю. В. и др., 1983] показало, что на первом этапе при умеренной активации ПОЛ появление гидроперекисей в липидном бислое мембран может привести к увеличению активности липид-белковых комплексов, обеспечивающих функционирование каналов ионной проницаемости. В дальнейшем, при прогрессировании ПОЛ и уменьшении количества ненасыщенных фосфолипидов в мембранах могут реализоваться по меньшей мере два обратимых явления: во-первых, указанные белки оказываются как бы вмороженными в матрицу и функциональная активность их снижается, во-вторых, в мембране формируются новые каналы ионной проводимости — перекисные кластеры, — которые делают мембрану более проницаемой для Са2+ и в меньшей степени для других катионов [Лебедев А. В. и др., 1980]. Наконец, еще большее нарастание процесса может привести к фрагментации мембраны, образованию сшивок между ее белками и фосфолипидами, окислению сульфгидрильных групп в активных центрах ферментов и, таким образом,— к необратимым повреждениям мембран и мембранно-связанных белков.
Оценивая эти данные, следует иметь в виду, что эффект стресса на сердце является по своему существу адренергическим и через систему адренорецепторов приводит к увеличенному вхождению Са2+ в кардиомиоциты. Данное явление играет роль в положительном инотропном эффекте катехоламинов и при умеренном стрессе оказывается транзиторным, так как благодаря нормальному функционированию мембранных механизмов ионного транспорта избыток Са2+ быстро удаляется из саркоплазмы. При чрезмерной по длительности стресс-реакции, повреждении мембран и катионных насосов удаление Са2+ из саркоплазмы может оказаться нарушенным, и в кардиомиоцитах может возрастать содержание Са2+. Действительно, сочетание методов гистохимии и электронной микроскопии позволило выявить в кардиомиоцитах при стрессе существенное увеличение количества Са2+ в субсарколеммальной области, саркоплазме и вокруг митохондрий [Меерсон Ф. 3., Самосудова Н. В. и др., 1983]. Это согласуется с известным фактом, что повреждение сердца большими дозами катехоламинов сопровождается увеличением накопления в миокарде Са2+.
Увеличение содержания Са2+ в кардиомиоцитах не является оригинальной принадлежностью повреждающего адренергического эффекта, а составляет общее звено патогенеза самых различных повреждений сердечной мышцы — от наследственных кардиопатий до недостаточности гипертрофированного сердца [Меерсон Ф. 3., Уголев А. А., 1980]. Данное изменение может способствовать чрезмерной активации гликолиза, разобщению окисления и фосфорилирования в митохондриях, нарушению процесса расслабления миофибрилл, вплоть до развития их контрактурного повреждения [Меерсон Ф. 3., 1984]. В соответствии с этим после эмоционально-болевого стрессорного воздействия обнаружена активация фосфорилазной системы — переход В-формы фосфорилазы в А-форму (Skinner J. E. et al., 1983], снижение резерва гликогена в миокарде на 1/3, разобщение окисления с фосфорилированием в митохондриях [Малышев В. В. и др., 1982 б] и снижение активности креатинфосфокиназы [Голубева Л. Ю., Меерсон Ф. 3., 1986]. В совокупности эти нарушения катионного и энергетического метаболизма закономерно приводят к целому комплексу нарушений электрической стабильности и сократительной функции сердца. Данный комплекс слагается из снижения порога фибрилляции сердца [Меерсон Ф. 3., Белкина А. М. и др., 1985] и аритмий [Lown В. et al., 1980], нарушения растяжимости сердечной мышцы — ее постстрессорной ригидности [Воронцова Е. Я., Пшенникова М. Г., Меерсон Ф. 3., 1982], депрессии силы, а также значительного повышения контрактурной реакции миокарда на избыток Са2+ и гипоксию. Важно, что эти нарушения метаболизма и функции, охватывающие миокард в целом, для большинства кардиомиоцитов оказываются обратимыми и постепенно исчезают в течение 3—4 сут. Лишь в ограниченных группах клеток они прогрессируют и приводят к разрушению структур. Соответственно морфологические изменения после перенесенного стресса у животных обычно носят очаговый характер, достигая максимума через 45—48 ч после стрессорного воздействия и проявляются при поляризационной микроскопии контрактурой групп мышечных клеток, которая в некоторых местах приводит к формированию микронекрозов, а затем фибробластических гранул и очагов кардиосклероза [Меерсон Ф. 3., 1984].
Представленная картина стрессорных повреждений миокарда, как указывалось, обладает глубоким сходством с изменениями, развивающимися под влиянием больших доз катехоламинов или их синтетического аналога изопротеренола и может быть предупреждена значительными дозами р-адреноблокатора пропранолола. Таким образом, повреждения сердца при стрессе носят адренергический характер.
Существенно, что все представленные выше компоненты адренергического повреждения сердца при стрессе, начиная от самой активации ПОЛ, увеличенного выхода ферментов из миокарда, нарушений активности ферментов катионного транспорта и кончая нарушениями электрической стабильности, сократительной
функции и структуры сердца, могут быть предупреждены или ограничены путем введения перед стрессорным воздействием синтетических или природных антиоксидантов. Этот результат, полученный в наших экспериментах при помощи ионола, веществ из класса оксипиридинов [Меерсон Ф. 3., Абдикалиев Н. А., Голубева Л. Ю., 1981 а], больших доз α-токоферола [Меерсон Ф. 3., Гибер Л. М. и др., 1977] говорит о том, что активация ПОЛ составляет важное, а возможно и ключевое звено патогенеза стрессорного повреждения сердца, а антиоксиданты, подавляющие ПОЛ в сердце и стресс-реакцию на центральном уровне [Меерсон Ф. 3., Малышев В. В. и др., 1982], являются фактором антистрессорной защиты. Это согласуется с установленным в последнее время фактом, что различные антиоксиданты: α-токоферол, цистеин и др. предупреждают повреждающий эффект и, в частности, аритмогенный эффект больших доз катехоламинов [Singal P. et al., 1985]. Такие наблюдения делают исключительно важным вопрос о механизме, через который избыток катехоламинов, возникающий пра стрессе, активирует свободнорадикальное окисление в миокарде.
В настоящее время можно выделить несколько предположительных механизмов адренергической активации свободнорадикального окисления.
1. Известно, что адренергический эффект сопряжен с большим расходом катехоламинов, поступающих в сердце и другие органы из симпатических терминалей и кровотока. В результате происходит значительное уменьшение содержания катехоламинов в мозге, сердце, надпочечниках и т. д. Распад катехоламинов реализуется, помимо прочего, через их автоокисление, которое сопровождается генерацией супероксиданионрадикала и, следовательно, может играть роль в активации ПОЛ [Bors U. et al., 1978]. В дальнейшем, после завершения непосредственного действия стрессорного фактора, происходит восстановление нормального содержания катехоламинов в сердце и других органах, в котором важную роль наряду с обратным захватом играет увеличение интенсивности ресинтеза. При этом образование катехоламинов из фенилаланина сопровождается генерацией свободных радикалов супероксида на каждой из трех стадий процесса [Bors U. et al., 1978]. Таким образом, активация распада и ресинтеза катехоламинов в сердце при стрессе может сама по себе индуцировать свободнорадикальное окисление и ПОЛ. Такое развитие событий при стрессе кажется весьма вероятным, однако при оценке его значения нужно учитывать, что β-блокатор пропранолол предупреждает адренергическую активацию ПОЛ в сердце при стрессе, а вместе с тем и повреждения миокарда [Пшенникова М. Г., Воронцова Е. Ф., 1983; Меерсон Ф. 3., 1984]. Это означает, что активация ПОЛ возникает скорее всего не как результат интенсификации метаболизма самих катехоламинов, а как следствие их действия на адренорецепторы. В связи с этим наибольшее значение приобретают механизмы адренергической активации ПОЛ, которые реализуются через систему адренорецептор — цАМФ.
2. Известно, что при стрессорном адренергическом эффекте катехоламины, связываясь с адренорецепторами, вызывают увеличение действующей концентрации цАМФ в миофибриллах, и, как следствие, рост потребления кислорода. Показано, что действие катехоламинов сопровождается усилением свободнорадикального окисления в мембранах митохондрий [Swaroop A., Ramasarma Т., 1981]. При анализе указанного факта следует учитывать, что в основе образования активных форм кислорода в митохондриях лежит «утечка» электронов из электронно-транспортной цепи [McCord J. M., Fridovich I., 1978], а сам процесс генерации свободных радикалов в митохондриях находится под контролем АТФ-синтетазной системы. Контроль этот осуществляется таким образом, что ингибирование АТФ-синтетазы и подавление ресинтеза АТФ прекращает свободнорадикальное окисление, а разобщение дыхания и фосфорилирования устраняет лимитирующую роль АТФ-синтетазы [Маршанский В. П., Новгородов С. А., 1981] и, по-видимому, может способствовать активации свободнорадикального окисления. При этом нужно учитывать, что большие дозы катехоламинов, подобные тем, которые действуют на сердце при стрессе, обладают разобщающим эффектом. Это выражается в том, что потребление кислорода сердцем возрастает больше, чем его работа [Challoner D., 1968], а концентрация креатинфосфата и АТФ снижается [Waldenstrom A. et al., 1978]. В плане нашего изложения существенно, что такое действие больших доз катехоламинов сопровождается, как недавно выяснилось, увеличением продуктов ПОЛ в мембранах митохондрий почти на 90% [Swaroop A., Ramasarma Т., 1981]. Таким образом, весьма вероятно, что увеличение генерации свободных радикалов митохондриями составляет один из факторов активации ПОЛ в миокарде при стрессе.
3. Другой возможный механизм адренергической активации ПОЛ связан с изменением в функционировании ферментной системы ксантиндегидрогеназы-ксантиноксидазы, ответственной за катаболизм конечных продуктов распада АТФ — ксантина и гипоксантина. В обычных условиях эта система представлена главным образом ксантиндегидрогеназой и не генерирует значимых количеств свободных радикалов. В последнее время возникло предположение, что при некоторых нарушениях клеточного гемостаза большая часть ксантиндегидрогеназы может под влиянием кальмодулинзависимых протеаз трансформироваться в ксантиноксидазу, активность которой сопряжена с генерацией супероксидного радикала (Chambers D. E. et al., 1985]. При адренергическом эффекте увеличение вхождения Са2+ в миокардиальные клетки хорошо известно, оно вызывает активацию протеаз и с большой долей вероятности может вызывать указанную трансформацию. Одновременно, при большом стрессорном адренергическом эффекте в миокарде реализуются два других фактора, необходимых для генерации свободных радикалов в ксантиноксидазной системе, а именно избыток кислорода и продуктов распада АТФ. Таким об-
разом, роль этого механизма в активации ПОЛ при стрессе следует считать вероятной.
4. При адренергическом эффекте катехоламины через цАМФ активируют фосфолипазу А2 [Higgins A., Blackburn К., 1984; Takasu N. et al., 1986], что может приводить к нарушению упорядоченности расположения липидов в клеточных мембранах. Такой хаотропный эффект может, как известно, активировать ПОЛ [Наtefi Y., Hanstein W., 1970]. Вероятно, что это явление действительно наблюдается при стрессорном повреждении сердечной мышцы, так как доказано, что ингибиторы фосфолипазы А2 могут предупреждать адренергические повреждения сердца [Higgins A., Blackburn К., 1984].
5. При стрессорном и других адренергических эффектах наблюдается опосредованное через адренорецепторы увеличенное вхождение Са2+ в кардиомиоциты. Доказано, что в физиологических концентрациях (от 10─7 до 5-10─5 М) Са2+ закономерно активирует ПОЛ в мембранах митохондрий и микросом сердечной мышцы [Каган В. Е., Савов В. М. и др., 1983], поэтому можно предположить, что кальциевая активация ПОЛ также составляет один измеханизмов общей активации ПОЛ в сердечной мышце. Это согласуется с фактом, что концентрации Са2+ выше 10─3 М, реально существующие в межклеточной жидкости, ингибируют ПОЛ [Каган В. Е., Архипенко Ю. В. и др., 1983] и это может служить причиной мелкоочаговых повреждений миокарда при стрессе.
Очевидно, что рассмотренные факторы не являютсяальтернативными и могут сочетаться между собой. В целом же молекулярный механизм стрессорной активации ПОЛ в сердечной мышце требует дальнейшего изучения.
Целесообразно подчеркнуть, что имеющиеся данные о ролиПОЛ в патогенезе стрессорного повреждения создают определенные перспективы для кардиологической клиники.
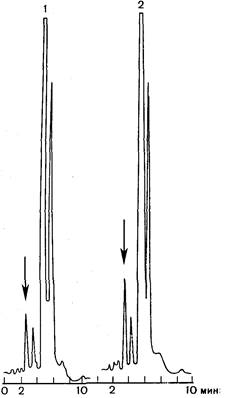
Во-первых, активация ПОЛ и защитное действие антиоксидантов при стрессе реализуются не только в сердце. Оба явления доказаны применительно к головному мозгу [Богданова Е. Д. и др., 1981], сетчатке глаза [Шведова А. А. и др., 1982, и др.], скелетным мышцам и т. д. Выяснилось, что активация ПОЛ при стрессе закономерно возникает у людей [Меерсон Ф. 3., Каган В. Е. и др., 1983]. Это было показано в специальных исследованиях, где активация ПОЛ в организме определялась путем измерения содержания пентана в выдыхаемом воздухе после напряженной операторской работы (рис. 24). Возможность неинвазивным путем количественно оценивать стрессорную активацию ПОЛ у человека может быть использована в функциональной диагностике для оценки стрессорных состояний и эффективности антиоксидантной защиты при заболеваниях сердца и других органов.
Во-вторых, описанные нарушения метаболизма, функции и структуры сердца наблюдаются после прекращения стрессорного воздействия и не только представляют собой реакцию на стресс, но приводят к стойким очаговым повреждениям, сохраняющимся
|
| Рис. 24. Хроматограммы воздуха, выдыхаемого людьми до стрессорного воздействия (1) и после него (2). Крайний слева пик хроматограмм — содержание пентана (стрелка). |
после того, как сам стресс миновал. Этот факт в совокупности с данными клиники о роли стресса в возникновении заболеваний кровообращения дает основание полагать, что именно такие повреждения, суммирующиеся от одного стрессорного эпизода к другому, могут играть роль в развитии первичного некоронарогенного кардиосклероза и хронической сердечной недостаточности, которые нередко возникают у людей, ранее не страдавших заболеваниями кровообращения. Таким образом, высокое содержание природных антиоксидантов в пище, а также специальная антиоксидантная защита при стрессорных ситуациях могут служить одним из факторов профилактики такого развития событий.
В-третьих, аритмогенный эффект стресса, как теперь доказано, может реализоваться независимо от ишемии или в сочетании с ней [Lown В. et al., 1980; Skinner J. E. 1985] и играет важную роль в механизме внезапной сердечной смерти.
Изложенное не сводится лишь к проблеме антиоксидантной защиты при стрессе, так как стресс часто выступает отнюдь не как самостоятельное явление, а как важное звено в патогенезесамых различных заболеваний и, в частности, в патогенезе ишемической болезни сердца, применительно к которой роль стресса показана в эксперименте и в клинике.
Факт активации ПОЛ в сердце под действием ишемии в настоящее время не вызывает сомнений. Такая активация показана при ишемии целого сердца, а также в зоне и вне зоны некроза при инфаркте миокарда (Коган А. X. и др., 1976; Меерсон Ф. 3., 1984]. Более того, ишемическая активация ПОЛ в сердце не уникальна, этот процесс является неспецифическим ответом на нарушение кровоснабжения многих органов и тканей: мозга, печени, почек, кишечника, конечностей и др. [Биленко М. В., 1978; Yoshida S. et al., 1982]. Установлено также, что предварительное введе-
ние антиоксидантных препаратов ограничивает подъем уровня ПОЛ в сердце, уменьшает зону некроза при инфаркте миокарда и предотвращает нарушение сократительной функции неишемизированных отделов, которые обычно повреждаются сопутствующим стрессом [Савов В. М. и др., 1985; Werns S. et al., 1985].
Вместе с тем факт ишемической активации ПОЛ на первый взгляд парадоксален, так как уменьшение или прекращение переноса кислорода к кардиомиоцитам отнюдь не должно способствовать развитию данного процесса. При выяснении причин этого кажущегося парадокса следует иметь в виду, что в случае острой ишемии и инфаркта миокарда в мышце сердца действует не один, а по меньшей мере 3 фактора, способных активировать ПОЛ: 1) ишемия, вызывающая поражения в зоне нарушенного кровотока; 2) стресс, повреждающий неишемизированные отделы сердца; 3) реоксигенация, действующая на границе раздела зон влияния стресса и ишемии.
Роль стресса и реоксигенации в повреждении сердца при инфаркте миокарда велика. Однако основным фактором повреждения при этом заболевании, триггером, способствующим реализации в мышце сердца всех последующих изменений, в том числе и таких как стрессорное повреждение и реоксигенация, является образование в миокарде очага ишемического повреждения. Активация ПОЛ под действием ишемии определяется формированием и интенсивным функционированием нескольких механизмов генерации свободных радикалов.
1. Механизм активации ПОЛ электронно-транспортными переносчиками. Он включается на ранних стадиях ишемии. В его основе лежит хорошо известный факт, что при движении электронов по дыхательной цепи всегда реализуется одна из двух потенциальных возможностей: электроны или доходят до конца цепи и под действием концевого фермента цитохромоксидазы переносятся на кислород с образованием воды, или же они передаются на кислород непосредственно с переносчиков цепи. Принципиальное различие между той и другой возможностями определяется тем, что при действии цитохромоксидазы радикалов не образуется, тогда как «утечка» электронов по пути следования к цитохромоксидазе приводит к образованию  Существенным является также то обстоятельство, что восстановление кислорода на цитохромоксидазе возможно лишь при бесперебойной и согласованной работе дыхательной цепи, т. е. в условиях нормального напряжения О2. При блокаде дыхательной цепи в ее конечном звене, т. е. при действии ишемической гипоксии первый — нерадикальный — путь функционирования дыхательной цепи подавляется; в этих условиях разгрузка дыхательной цепи от постоянно пополняющих ее электронов происходит за счет активации второго, радикалообразующего пути. Изложенная концепция подтверждается многочисленными экспериментальными фактами.
Существенным является также то обстоятельство, что восстановление кислорода на цитохромоксидазе возможно лишь при бесперебойной и согласованной работе дыхательной цепи, т. е. в условиях нормального напряжения О2. При блокаде дыхательной цепи в ее конечном звене, т. е. при действии ишемической гипоксии первый — нерадикальный — путь функционирования дыхательной цепи подавляется; в этих условиях разгрузка дыхательной цепи от постоянно пополняющих ее электронов происходит за счет активации второго, радикалообразующего пути. Изложенная концепция подтверждается многочисленными экспериментальными фактами.
Так, показана возможность активной генерации  восстановленными электронно-транспортными переносчиками НАД, НАДФ, ФАД, KoQ [Fridovich I., 1980; Rao P. et al., 1983]. Блокада пути переноса электронов антимицином А — средством, препятствующим поступлению ё к цитохромоксидазе,— т. е. моделирование ситуации, сходной с ишемической, приводит к резкому увеличению образования
восстановленными электронно-транспортными переносчиками НАД, НАДФ, ФАД, KoQ [Fridovich I., 1980; Rao P. et al., 1983]. Блокада пути переноса электронов антимицином А — средством, препятствующим поступлению ё к цитохромоксидазе,— т. е. моделирование ситуации, сходной с ишемической, приводит к резкому увеличению образования 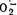 и Н2О2 [Ramasarma Т., 1982]. К такому же результату ведет даже кратковременное помещение митохондрий сердца в условия аноксии. Уровень генерации
и Н2О2 [Ramasarma Т., 1982]. К такому же результату ведет даже кратковременное помещение митохондрий сердца в условия аноксии. Уровень генерации 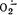 при этом возрастает в 7 раз [Леднев А. Н., Рууге Э. К., 1985]. Тем не менее в условиях дефицита кислорода генерация активных форм О2 и интенсификация ПОЛ лимитированы. Значительно большее образование активных форм О2 и продуктов ПОЛ происходит в условиях, когда реализовавшееся во время ишемии накопление восстановленных переносчиков цепи — доноров электронов — сочетается с достаточно высоким напряжением кислорода, т. е. когда вслед за достаточно длительной ишемией и аноксией возникает избыток О2— акцептора электронов.
при этом возрастает в 7 раз [Леднев А. Н., Рууге Э. К., 1985]. Тем не менее в условиях дефицита кислорода генерация активных форм О2 и интенсификация ПОЛ лимитированы. Значительно большее образование активных форм О2 и продуктов ПОЛ происходит в условиях, когда реализовавшееся во время ишемии накопление восстановленных переносчиков цепи — доноров электронов — сочетается с достаточно высоким напряжением кислорода, т. е. когда вслед за достаточно длительной ишемией и аноксией возникает избыток О2— акцептора электронов.
Это состояние, обозначаемое как реоксигенация, характеризуется значительным накоплением вторичных продуктов ПОЛ [Guarnieri G. et al., 1980] и является фактором, эффективно разрушающим клеточные мембраны. При нем быстро реализуется повреждение кардиомиоцитов, проявляющееся увеличенным освобождением в перфузат ферментов КФК, ЛДГ, разрушением сарколеммы, СПГ, набуханием и разрыванием митохондрий, а также появлением в саркоплазме и митохондриях большого количества гранул фосфата кальция и полной контрактурой миофибрилл, при которой диски I на электронограммах не просматриваются [Hearse D. et al., 1980].
Вклад реоксигенации в активацию ПОЛ и повреждение сердца при инфаркте миокарда определяется тем, что зона первичной ишемии всегда больше, чем зона последующего некроза, так как часть первично ишемизированной ткани получает в дальнейшем достаточно эффективное кровоснабжение вследствие аденозиновой гиперемии и раскрытия коллатералей. Такое восстановление кровоснабжения, естественно, должно сопровождаться явлениями реоксигенации. Повреждающее действие ее в этом случа
|
|
|
