 |


Межнуклеотидная связь в РНК
|
|
|
|
Более сложным оказался вопрос о природе межнуклеотидной связи в РНК. Уже на первых этапах изучения строения РНК был установлен факт чрезвычайной неустойчивости се при щелочном гидролизе. Основными продуктами щелочного гидролиза РНК являются рибонуклсозид-2'- и рибонуклеозид-З'-фосфаты, образующиеся практически в равных количествах.

Рибонуклеозид-5'-фосфаты при этом не образуются. Эти данные не укладывались в представления о фосфодиэфирной природе межнуклеотидной связи в РНК и требовали всестороннего изучения. Очень важную роль в таком исследовании, которое выполнили в начале 50-х гг. Тодд с сотрудниками, сыграли синтетические алкиловые эфиры рибонуклеотидов, которые были получены специально, чтобы промоделировать тот или иной тип фосфодиэфирной связи.
Полученные школой Тодда данные о механизмах превращения алкиловых эфиров рибонуклеотидов в щелочной среде позволили предположить, что в РНК, так же как и в ДНК, межнуклеотидная связь осуществляется фосфатной группой и 3'- и 5'-гидроксильными группами углеводных остатков. Подобная связь в РНК должна очень легко расщепляться в щелочной среде, так как соседняя 2'-гидроксильная группа должна катализировать этот процесс при рН>10, когда начинается ионизация гидроксильных групп рибозы. Очень важно подчеркнуть, что промежуточными соединениями при щелочном расщеплении должны быть все четыре рибонуклеозид-2',З'-циклофосфата, а конечными — образующиеся при их гидролизе рибонуклеозид-3'-фосфаты и рибонуклеозид-2'-фосфаты (четыре пары изомеров).
Данные щелочного гидролиза ограничили количество возможных для РНК типов межнуклеотидных связей, но не прояснили вопроса о том, как построен этот полимер.
|
|
|
Более точные сведения о типе межнуклеотидной связи в РНК, как и в случае ДНК, были получены с помощью ферментативного гидролиза.
Гидролиз РНК с использованием ФДЭ змеиного яда, протекающий до рибонуклеозид-5'-фосфатов, подтвердил уже прямым путем предположение об участии 5'-гидроксильных групп в образовании фосфодиэфирной связи между мономерными звеньями. Позднее это было окончательно установлено в результате открытия фосфоролиза РНК в присутствии фермента полинуклеотидфосфорилазы (ПНФаза), приводящего к образованию рибонуклеозид-5'-пирофосфатов:
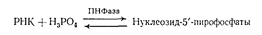
Таким образом, оставалось выяснить природу второй гидроксильной группы, участвующей в образовании межнуклеотидной связи. Частично решить эту задачу помог еще один фермент, который использовался для направленного расщепления РНК, — пиримидиловая рибонуклеаза (РНаза).
Ранее было показано, что этот фермент расщепляет только алкиловые эфиры пиримидиновых рибонуклеозид-3'-фосфатов до рибонук-леозид-3'-фосфатов (через промежуточный рибонуклеозид-2',З'-циклофосфат). Оказалось, что аналогичным образом этот фермент действует и на РНК. В экспериментах с любыми образцами очищенной РНК было обнаружено, что количество фосфорной кислоты, которая образуется при обработке полимера последовательно пиримидиловой РНазой и фосфомоноэстеразой (ФМЭ), а также количество иодной кислоты, затрачиваемой на последующее окисление, эквивалентно количеству остатков пиримидинов в данном образце РНК. Это говорило в пользу того, что по крайней мере пиримидиновые нуклеотиды в РНК связаны с соседними нуклеотидами только посредством 3'—5'-межнук-леотидной связи. Этот вывод подтверждают данные щелочной обработки ферментативных гидролизатов РНК, полученных после действия на нее РНазы: в щелочной среде миграция фосфатного остатка в рибонуклеозид-З'- и -2'-фосфатах невозможна, и наличие в соответствующих гидролизатах только пиримидиновых рибонуклеозид-З'-фосфатов делает очевидным 3'—5'-тип межнуклеотидной связи для пиримидиновых нуклеотидов.
|
|
|
МАТРИЧНЫЙ СИНТЕЗ ДНК
Способность клеток поддерживать высокую упорядоченность своей организации зависит от генетической информации, которая сохраняется в форме дезоксирибонуклеиновой кислоты (ДНК). ДНК - это вещество, из которого состоят гены. Размножение живых организмов, передача наследственных свойств из поколения в поколение и развитие многоклеточного организма из оплодотворенной яйцеклетки возможны потому, что ДНК способна к самовоспроизведению. Сам процесс самовоспроизведения ДНК называется репликацией. Иногда используют также название-синоним - редупликация.
Как известно, генетическая информация записана в цепи ДНК в виде последовательности нуклеотидных остатков, содержащих одно из четырех гетероциклических оснований: аденин (A), гуанин (G), цитозин (C) и тимин (T). Предложенная Дж. Уотсоном и Ф. Криком в 1953 году модель строения ДНК в форме регулярной двойной спирали сразу же позволила понять принцип удвоения ДНК. Информационное содержание обеих цепей ДНК идентично, так как каждая из них содержит последовательность нуклеотидов, строго соответствующую последовательности другой цепи. Это соответствие достигается благодаря наличию водородных связей между направленными навстречу друг другу основаниями двух цепей - попарно G и C или A и T. Описывая это свойство двойной спирали, молекулярные биологи говорят, что цепи ДНК комплементарны за счет образования уотсон-криковских пар GРC и AРT.
Поскольку две цепи имеют противоположную направленность, их называют антипараллельными. Легко представить, что удвоение ДНК происходит вследствие того, что цепи расходятся, а потом каждая цепь служит матрицей, на которой собирается комплементарная ей новая цепь ДНК. В результате образуются две дочерние, двуспиральные, неотличимые по строению от родительской ДНК молекулы. Каждая из них состоит из одной цепи исходной родительской молекулы ДНК и одной вновь синтезированной цепи. Такой механизм репликации ДНК, при котором от одного поколения к другому передается одна из двух цепей, составляющих родительскую молекулу ДНК, получил название полуконсервативного и был экспериментально доказан в 1958 году М. Мезельсоном и Ф. Сталь.
|
|
|
Кроме того, ситезу ДНК характерны такие свойства, как антипараллельность и униполярность. Каждая цепь ДНК имеет определенную ориентацию. Один конец несет гидроксильную группу (ОН), присоединенную к 3'-углероду в сахаре дезоксирибозе, на другом конце цепи находится остаток фосфорной кислоты в 5'-положении сахара. Две комплементарные цепи в молекуле ДНК ориентированы в противоположных направлениях - антипараллельно (при параллельной ориентации напротив 3'-конца одной цепи находился бы 3'-конец другой). Ферменты, синтезирующие новые нити ДНК, называемые ДНК-полимеразами, могут передвигаться вдоль матричных цепей лишь в одном направлении - от их 3'-концов к 5'-концам. При этом синтез комплементарных нитей всегда ведется в 5' 3' направлении, то есть униполярно. Поэтому в процессе репликации одновременный синтез новых цепей идет антипараллельно.
ДНК-полимеразы могут давать "задний ход", то есть двигаться в направлении 3' 5'. В том случае, когда последнее добавленное при синтезе нуклеотидное звено оказалось некомплементарным нуклеотиду матричной цепи, оно будет замещено комплементарным нуклеотидом. Отщепив "неправильный" нуклеотид, ДНК-полимераза продолжает синтез в 5' 3' направлении. Такая способность к исправлению ошибок получила название корректорской функции фермента.
ДНК-полимеразы
В 1957 году А. Корнберг обнаружил у кишечной палочки фермент, катализирующий процесс полимеризации ДНК из нуклеотидов; он был назван ДНК-полимеразой. Затем ДНК-полимеразы выявили и в других организмах. Было показано, что субстратами всех этих ферментов служат дезоксирибонуклеозидтрифосфаты (дНТФ), полимеризующиеся на одноцепочной ДНК-матрице. ДНК-полимеразы последовательно наращивают одноцепочную цепь ДНК, шаг за шагом присоединяя к ней следующие звенья в направлении от 5-' к 3'-концу, причем выбор очередного дНТФ диктуется матрицей. Присоединение каждого нового нуклеотидного остатка к 3'-концу растущей цепи сопровождается гидролизом богатой энергией связи между первым и вторым фосфатными остатками в дНТФ и отщеплением пирофосфата, что делает реакцию в целом энергетически выгодной.
В клетках обычно присутствует несколько типов ДНК-полимераз, выполняющих различные функции и имеющих разное строение. Они могут быть построены из различного количества белковых цепей (субъединиц), от одной до десятков, однако все они работают на любых последовательностях нуклеотидов матрицы. Задача этих ферментов - сделать точную копию каждой матрицы.
|
|
|
