 |


Принципы экологической классификации организмов
|
|
|
|
Современная систематика растений и животных строится на основе единственного главного критерия — степени родства организмов. При этом внешние особенности видов, относимых к одной группе, часто могут сильно различаться. Так, например, паразит крабов саккулина, напоминающая бесформенный, набитый половыми продуктами мешок с сильно разветвленной в теле хозяина сетью тяжей, внешне совершенно не похожа на сидячих, обладающих раковинам» морских желудей и морских уточек, хотя все они относятся к одному отряду усоногих раков. О родстве этих видов говорит глубокое внутреннее сходство, прослеживаемое на первых этапах развития особей.
В экологии разнообразие и разноплановость способов и путей адаптации к среде создают необходимость множественных классификаций. Используя какой-либо единственный критерий, нельзя отразить все стороны приспособленности организмов к среде. Экологические классификации отражают сходство, возникающее у представителей самых разных групп, если они используют сходные пути адаптации. Например, если мы классифицируем животных по способам движения, то в экологическую группу видов, передвигающихся в воде реактивным путем,, попадут такие- разные но систематическому положению животные, как медузы, головоногие моллюски, некоторые инфузории и жгутиковые, личинки ряда стрекоз и др. В основу экологических классификаций могут быть положены самые разнообразные критерии,: способы питания, передвижения, отношение к температуре, влажности, -солености среды, давлению и т. п. Разделение всех организмов на эврибионтных и стенобионтных по широте диапазона приспособлений к среде представляет пример простейшей экологической классификации. Другой пример—разделение организмов на группы по характеру питания.
|
|
|
Автотрафы — это организмы, использующие в качестве источника для построения своего тела неорганические соединения. Гетеротрофы — все живые существа, нуждающиеся в пище органического происхождения. В свою очередь, автотрофы делятся на фотографов и хемотрофов. Первые для синтеза органических молекул используют энергию солнечного света, вторые — энергию химических связей. Гетеротрофов делят на сапрофитов, использующих растворы простых органических соединений, и голозоев. Голозои обладают сложным комплексом пищеварительных ферментов и могут употреблять в пищу сложные органические соединения, разлагая их на более простые составные компоненты. Голозои делятся на сапрофагов (питаются мертвыми растительными остатками),, фитофагов (потребителей живых растений), зоофагов (нуждающихся в живой пище) и некрофагов (трупоядных животных). В свою очередь, каждую из этих групп можно подразделить на более мелкие, имеющие свою специфику в характере питания.
Иначе можно построить классификацию по способу добывания пищи. Среди животных выявляются, например, такие группы, как фильтраторы (мелкие рачки, беззубка, кит и др.), пасущиеся формы (копытные, жуки-листоеды), собиратели (дятлы, кроты, землеройки, куриные), охотники на движущуюся добычу (вояки, львы, мухи-ктыри и т. п.) и целый ряд других групп. Так, несмотря на большое несходство в организации, одинаковый способ овладения добычей приводит у львов и мух-ктырей к ряду аналогий в их охотничьих повадках и общих чертах строения: поджарости, тела, сильному развитию мускулатуры, способности развивать кратковременно большую скорость и т. п.
Экологические классификации помогают выявлять возможные в природе пути приспособления организмов к среде.
ЖИЗНЕННЫЕ ФОРМЫ ОРГАНИЗМОВ
|
|
|
Среди приспособлений животных и растений к среде немаловажную роль играют морфологические адаптации, т. е. такие особенности внешнего строения, которые способствуют выживанию и успешной жизнедеятельности организмов в обычных для них условиях. «Целесообразность» строения живых существ долго не могла получить материалистического объяснения. Ч. Дарвин, выдвигая теорию естественного отбора как главного фактора эволюции, обратил внимание па то, что результатом естественного отбора может быть не только нарастание различий между близкими видами, но выработка у неродственных форм внешнего сходства, если эти виды ведут сходный образ жизни в близких условиях среды. Этот процесс получил название конвергенции. Конвергенция признаков у разных форм в наибольшей мере затрагивает те органы, которые находятся в непосредственном соприкосновении с внешней средой. Внутренние черты строения организмов, их общий план строения остаются при этом неизменными, отражая родство и происхождение видов.
Формообразующая роль факторов среды, т. е. влияние их на морфологию организмов, наглядно выступает при изучении роли влажности, температуры, движения воды и воздуха, плотности среды, объема пригодного для жизни пространства и др. Чем жестче физические условия среды, тем ограниченнее пути приспособления к ней. Одинаковые принципы освоения среды ведут к выработке сходных морфологических адаптации у разных видов, даже сильно различающихся по плану своего строения.
На форму быстро движущегося в жидкости тела накладывают жесткие ограничения законы физики. На него действует лобовое сопротивление, которое зависит от природы жидкости, скорости движения, площади проекции тела перпендикулярно направлению движения и длины тела в направлении потока. Для жидкости с такой вязкостью и плотностью, как вода, лобовое сопротивление для тел одинакового объема минимально, если отношение длины к наибольшему диаметру равно примерно 4,5. Хорошие пловцы среди животных обладают именно такими пропорциями, хотя достигают этого разными средствами. Плывущие кальмары так складывают щупальца, что тело становится торпедообразным, с оптимальным соотношением длины и диаметра, их скорость достигает 41 км/ч. У рыб большое значение имеет форма хвостового плавника; тюлени соединяют и вытягивают задние ласты. По пропорциям тела можно судить о максимально возможной скорости животного в воде. Дельфин-белобочка может плыть со скоростью 44 км/ч, тогда как тюлени, пропорции которых менее оптимальны,— не более 20 км/ч. Обтекаемость тела, таким образом,— необходимый путь приспособления к быстрому движению в воде.
|
|
|
Среди самых разнообразных в систематическом отношении планктонных организмов (одноклеточных водорослей, кишечнополостных, ракообразных, моллюсков, червей, личинок разных групп беспозвоночных, пелагической икры рыб и т. д.) мы встречаем лишь три описанных выше способа приспособления к парению: 1) уменьшение объема тела, 2) развитие разнообразных выростов, 3) увеличение содержания в теле воды, жиров и газообразных продуктов. Во всех случаях достигается один эффект: уменьшение отношения массы тела к его площади, что обеспечивает уравновешивание сил тяжести и трения о воду.
Таким образом, общее число возможных морфологических приспособлений к одной и той же среде ограничено.
Морфологический тип приспособления животного или растения к основным факторам местообитания и определенному образу жизни называют жизненной формой организма.
Жизненные формы растений
Жизненные формы сначала были выделены у растений. Еще Теофраст классифицировал растения, выделяя среди них деревья, кустарники, полукустарники и травы.
Термин «жизненная форма» был впервые употреблен в 1884 г. датским ботаником Е. Вармингом, который понимал под этим «форму, в которой вегетативное тело растения находится в гармонии с внешней средой в течение всей его жизни, от колыбели до гроба, от семени до отмирания».
Как синонимы или близкие понятия в экологии растений употребляются термины «экобиоморфа», «биологический тип», «форма роста», «эпиморфа».
Классификация жизненных форм. Для классификации жизненных форм датский ботаник К- Раункиер выбрал только один признак, но имеющий большое приспособительное значение: положение почек или верхушек побегов в течение неблагоприятного времени года по отношению к поверхности почвы и снегового покрова. К- Раункиер разработал классификацию жизненных форм вначале для растений Средней Европы, но затем распространил ее и на растения тропического пояса, таким образом, она стала универсальной.
|
|
|
Все растения К. Раупкиер разделил на пять типов.
I. Фанерофиты (Рп) — почки возобновления открытые или закрытые, расположены над поверхностью почвы высоко (выше 30 см). Подразделяются на 15 подтипов по консистенции стебля, по высоте растений, по ритму развития листвы, по защищенности почек.
II. X а м е ф и т ы (СЬ) — почки возобновления у поверхности почвы или не выше 20—30 см. Подразделяются на 4 подтипа.
III. Г е м и к р и п т о ф и т ы (НК) — почки возобновления на поверхности почвы или в самом поверхностном ее слое, покрытом часто подстилкой. Включают 3 подтипа и более мелкие подразделения.
IV. Криптофиты (К)—почки возобновления скрыты в почве (геофиты) или под водой (гелофиты и гидрофиты). Подразделяются на 7 подтипов.
V. Терофиты (ТЬ)—возобновление после неблагоприятного времени года только семенами.
Подразделение на подтипы основано на использовании морфо-биологических признаков (характер и расположение побегов, защищенность почек и др.).
К. Раупкиер считал, что жизненные формы складываются исторически как результат приспособления растений к климатиче ским условиям среды. Процентное распределение видов по жизненным формам в растительных сообществах на изучаемой территории он назвал биологическим спектром. Для разных зон и стран были составлены биологические спектры, которые могли служить индикаторами климата. Так, жаркий и влажный климат тропиков был назван климатом фамерофитов, а континентальный климат уморенного пояса—климатом гемикриптофитов.
 Однако типы жизненных форм К. Раункиера слишком обширны и неоднородны. Хамефиты, например, включают растения с разным отношением к климату. Их много, с одной стороны, в тундрах, а с другой — в пустынях. Тем не менее классификация жизненных форм растений по Раункиеру остается популярной и продолжает модифицироваться.
Однако типы жизненных форм К. Раункиера слишком обширны и неоднородны. Хамефиты, например, включают растения с разным отношением к климату. Их много, с одной стороны, в тундрах, а с другой — в пустынях. Тем не менее классификация жизненных форм растений по Раункиеру остается популярной и продолжает модифицироваться.
В архитектонике жизненных форм растений решающее значение имеют вегетативные органы. Согласно определению, данному И. Г. Серебряковым, жизненная форма —это своеобразный общий облик (габитус) определенной группы растений, сложившийся в их онтогенезе в результате роста и развития в определенных условиях среды. Этот габитус исторически возникает как выражение приспособленности растений к конкретным почвенно-климатическим условиям, как благоприятным, так и неблагоприятным. Именно система вегетативных органов создает жизненную форму, будучи необходимой в каждый момент жизни данного растения. Таким образом, жизненная форма — категория морфологическая и экологическая.
|
|
|
Обычно в классификации жизненных форм имеют в виду взрослую генеративную нормально развитую особь вида. Однако в онтогенезе растения происходит неоднократная смена жизненной формы. Ювелирные растения многих поликарпических трав стержнекорневые. С возрастом они могут утрачивать стержневую корневую систему и становятся кистекорневыми (купальница европейская, синец луговой) или короткокорневищными (вероника колосистая, василек сибирский).
 Разнообразие жизненных форм. Наиболее разработанной современной классификацией жизненных форм покрытосеменных и хвойных на основе -эколого-морфологических признаков является классификация, предложенная И. Г. Серебряковым. В ней использована большая совокупность признаков в соподчиненной системы приняты следующие единицы классификации: отделы, типы, классы, подклассы, группы, подгруппы, секции жизненных форм и собственно жизненные формы. Собственно жизненная форма может быть уподоблена виду в систематике и является основной единицей экологической системы растений.
Разнообразие жизненных форм. Наиболее разработанной современной классификацией жизненных форм покрытосеменных и хвойных на основе -эколого-морфологических признаков является классификация, предложенная И. Г. Серебряковым. В ней использована большая совокупность признаков в соподчиненной системы приняты следующие единицы классификации: отделы, типы, классы, подклассы, группы, подгруппы, секции жизненных форм и собственно жизненные формы. Собственно жизненная форма может быть уподоблена виду в систематике и является основной единицей экологической системы растений.
 Выделение отделов основано на структуре надземных осей (древесные, полудревесные и травянистые растения), типов — на относительной длительности жизни надземных осей (в отделе древесных) или на длительности жизни растений в целом (в отделе надземных травянистых растений).
Выделение отделов основано на структуре надземных осей (древесные, полудревесные и травянистые растения), типов — на относительной длительности жизни надземных осей (в отделе древесных) или на длительности жизни растений в целом (в отделе надземных травянистых растений).
Классы в пределах типов выделяют на основе особенностей структуры побегов (например, растения со стелющимися, лиановидными и другими побегами). Более мелкие таксономические единицы — по более частным признакам.
Эволюция видов и освоение ими новых экологических ниш привели к появлению огромного разнообразия жизненных форм.
Споровые растения — первенцы наземной растительности сохраняли сходство со своими предками — водорослями. Это были растения, близкие к травянистым, так как они имели небольшие размеры и свойственную травам совокупность анатомических и морфологических черт. Позднее возникли древовидные формы — лепидодендроны, сигиллярии, каламиты, древовидные папоротники. Вместе с ними, по-видимому, всегда существовали и травянистые папоротниковидные, отличавшиеся большим разнообразием жизненных форм. Потомки их дожили до наших дней, а значительная часть древовидных форм вымерла. Голосеменные, наоборот, представлены преимущественно древесными формами.
Очень разнообразны жизненные формы у господствующих ныне покрытосеменных, особенно в отделах древесных растений и наземных трав.
Сходные формы растений возникли конвергентно в разных систематических группах. Например, в условиях аридного климата пустынь одна и та же жизненная форма стеблевых суккулентов у кактусов (неотропис) и у молочаев (палеотропис). Одну жизненную форму могут иметь как близкородственные виды (например, у манжеток), так и виды, далеко отстоящие друг от друга. В то же время у одного вида на протяжении его ареала в разных географических и экологических условиях, в разных ценопопуляциях (совокупность особей данного вида в пределах одного фитоценоза) или даже в одной ценопопуляцпи встречаются взрослые особи, представляющие собой различные жизненные формы.
Пространственная дифференцировка вида у растений очень сложная, поэтому изучение ценопопуляций привлекает особое внимание.
В североказахстанских степях в ценопопуляциях подмаренника русского встречаются две жизненные формы: длиннокорневищно-стержпекорневая и дерновинно-стержнекорневая. Первая преобладает на более рыхлых почвах, вторая — на более плотных. В ценопопуляцпях луговика извилистого встречаются растения с компактной дерновиной и длинностолоновые полицентрические особи с удаленными друг от друга парциальными кустами на десятки сантиметров. В искусственных фитоценозах газонного типа одуванчик лекарственный встречается как стержнекорневое и корнеотпрысковое растение.
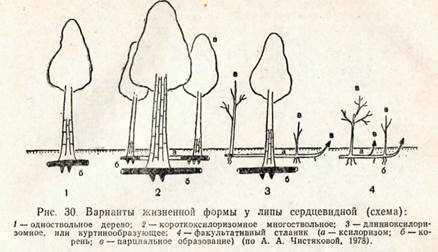
Многие виды деревьев на границах ареала образуют кустарниковые, нередко стелющиеся формы, например ель обыкновенная на Крайнем Сопоре, ель сибирская — на Южном Урале и в Хибинах. Липа сердцелистная и клен полевой в пределах ареала встречаются в виде одноствольного дерева, немногоствольного дерева (дерево-куст), в виде куртины (куртинообразующее дерево) и кустовидного стланика. Добавочные стволы у компактного многоствольного дерева и в куртине связаны с основным стволом одревесневшими корневищами длиной от нескольких сантиметров до 4—5 м. Компактные древовидные жизненные формы липы преобладают па юге и в центре ареала (па Украине, в Тульской и Пензенской областях), а стланиковые — близ северо-восточной границы на Среднем Урале. В пределах одного фитоценоза могут встречаться все жизненные формы липы. Кустовидный стланик— это форма угнетенных растений липы, она встречается при сильном затенении, а также приурочена к склонам, днищам оврагов, влажным местообитаниям. Такие экземпляры не цветут, имеют высоту до 4 м, создают подлесок. Сходные варианты жизненных форм встречаются и у черемухи обыкновенной. Стланиковые формы липы, черемухи и клопа полового при улучшении условий произрастания переходят в кустовидные формы или становятся куртинообразующими доревьями, и молодое одноствольное дерево может стать КурТИНООбразуЮЩИМ.
 СтлаНИКОВЫе формы могут быть и у многих кустарников — жимолости лесной, бсресклета бородавчатого и европейского, свидины. Лимонник китайский в лесах Дальнего Востока в разных экологических условиях растет то как лиана, то как кустарник.
СтлаНИКОВЫе формы могут быть и у многих кустарников — жимолости лесной, бсресклета бородавчатого и европейского, свидины. Лимонник китайский в лесах Дальнего Востока в разных экологических условиях растет то как лиана, то как кустарник.
Наиболее разнообразны жизненные формы деревьев в растительных сообществах тропического пояса. Габитус тропических деревьев определяется зачастую не только характером стволов и крон, но и корневыми системами, поэтому последние служат важным признаком при классификации жизненных форм деревьев.
Широко признано, что значительная часть родов и семейств покрытосеменных в процессе эволюции претерпела соматическую редукцию. От крупных деревьев с хорошо развитым стволом и сильно разветвленной кроной произошли немногоствольные деревья, а затем кустарники, кустарнички и различные травы. Но некоторые семейства с самого начала, по-видимому, состояли из травянистых видов, от которых возникли наиболее специализированные древесные формы, например бамбуки в семействе злаков.
Травянистые растения, имеющие более короткую длительность жизни надземных осей, разнообразные ритмы сезонного развития, разный характер подземных органов, часто вегетативно подвижные или обладающие большой семенной продуктивностью, лучше приспособлены к освоению самых различных местообитаний, иногда с очень суровыми условиями. Поэтому разнообразие жизненных форм у наземных травянистых растений необыкновенно велико.
В сходных условиях как среди древесных, так и среди травянистых растений конвергентно возникли лиановидные, суккулентные, стелющиеся, подушковидные формы, составляющие параллельные ряды. Например, подушковидные формы древесные и травянистые часто встречаются в условиях хорошего освещения, но при низких температурах воздуха и почвы, при крайней сухости почвы и низкой влажности воздуха, при частых и сильных ветрах. Обычны они в высокогорьях, тундрах, пустынях, на субантарктических островах и побережьях и в других местах с подобным комплексом условий.
Жизненные формы животных
Классификации жизненных форм животных, как и растений, весьма разнообразны в зависимости от принципов, которые кладут в их основу. На морфологии млекопитающих, по-видимому, в наибольшей степени сказывается характер их передвижения в разных средах. А. Н. Формозов выделил на этом основании среди зверей следующие адаптивные типы: 1) наземные формы, 2) подземные (землерои), 3) древесные, 4) воздушные и 5) водные. Между типами есть переходы.

 В пределах каждой группы особенности поступательного движения и образа жизни формируют более специфические приспособительные формы. Так, наземные млекопитающие передвигаются в основном посредством ходьбы, бега и прыжков, что проявляется в их внешнем облике. Например, прыгающие животные: кенгуру, тушканчики, прыгунчики, кенгуровые крысы — отличаются компактным телом с удлиненными задними конечностями и значительно укороченными передними. Очень сильно развиты мышцы — разгибатели спины, увеличивающие мощность толчка. Длинный хвост играет роль балансира и руля, позволяющего резко изменять направление движения. У сидящих животных он служит дополнительной точкой опоры. Сходный тип передвижения и внешним облик имели и некоторые вымершие динозавры мезозоя. Естественно, что, кроме общих адаптивных признаков, каждый вид отличается деталями внешнего строения в зависимости от экологической специфики.
В пределах каждой группы особенности поступательного движения и образа жизни формируют более специфические приспособительные формы. Так, наземные млекопитающие передвигаются в основном посредством ходьбы, бега и прыжков, что проявляется в их внешнем облике. Например, прыгающие животные: кенгуру, тушканчики, прыгунчики, кенгуровые крысы — отличаются компактным телом с удлиненными задними конечностями и значительно укороченными передними. Очень сильно развиты мышцы — разгибатели спины, увеличивающие мощность толчка. Длинный хвост играет роль балансира и руля, позволяющего резко изменять направление движения. У сидящих животных он служит дополнительной точкой опоры. Сходный тип передвижения и внешним облик имели и некоторые вымершие динозавры мезозоя. Естественно, что, кроме общих адаптивных признаков, каждый вид отличается деталями внешнего строения в зависимости от экологической специфики.
Жизненные формы отчетливо выделяются в пределах любой крупной таксономической группы животных, характеризующейся экологическим разнообразием видов.
Во внешнем облике птиц в наибольшей мере проявляется приуроченность их к определенным типам местообитаний и характер передвижения при добывании пищи. В связи с этим различают птиц: 1) древесной растительности, 2) открытых пространств суши, 3) болот и отмелей, 4) водных пространств. В каждой из указанных групп выделяют специфические формы: а) добывающие пищу посредством лазания (многие голуби, попугаи, кукушки, дятловые, воробьиные); б) добывающие пищу в полете (в лесах — совы, козодои и др., на открытых пространствах — ржанковые, длиннокрылые, над водой — трубконосые и т. п.); в) кормящиеся при передвижении по земле (лесные — большинство куриных, казуары, киви и др., на открытых пространствах — страусы, нанду,--журавлиные, на болотах и отмелях — голенастые, фламинго, некоторые воробьиные); г) добывающие пищу с помощью плавания и ныряния (пингвины, гагары, поганки, ряд трубконосых, большинство веслоногих и гусиных и др.).
 Жизненные формы таких мелких почвенных членистоногих, как коллемболы, выделяют па основе приуроченности их к определенным слоям почвенного профиля, так как в почве с глубиной резко меняется весь комплекс условий обитания: размеры полостей, освещенность, режим температуры и влажности и т. п.
Жизненные формы таких мелких почвенных членистоногих, как коллемболы, выделяют па основе приуроченности их к определенным слоям почвенного профиля, так как в почве с глубиной резко меняется весь комплекс условий обитания: размеры полостей, освещенность, режим температуры и влажности и т. п.
Атмобионты — виды, населяющие верхние слои подстилки и способные подниматься на нижние части растений, характеризуются более крупными размерами, полностью развитыми глазами, относительно длинными придатками (антеннами, ногами и прыгательной вилкой), хорошо выраженной пигментацией. Эуздафические виды — обитатели минеральных тонкопорозных слоев почвы— отличаются мелкими размерами, сильным укорочением придатков, редукцией вилки, полным отсутствием глаз и пигмента. Гемиэдафические формы характеризуются промежуточными чертами: частичной редукцией глаз, рассеянным пигментом, укороченной вилкой и др.
Среди саранчовых по форме (конфигурация тела, структура головы, строение конечностей, летательного аппарата, тип покровительственной окраски) хорошо различаются тамнобионты — обитатели кустарником и деревьев; населяющие травянистый ярус хортобианты; жители падпочтчшого слоя органических остатков герпвтобионты; обитвтели открытых участков грунта: эремобионты — на поверхности плотных глинистых почв, псаммобионты — на песках и петробионты— на каменистых участках с разреженной растительностыо.
Сходные жизненные формы встречаются в сходных условиях жизни в разных зоогеографических областях и на разных материках (например, тушканчики Евразии, кенгуровые крысы Австралии, прыгунчики Африки). Жизненная форма наглядно свидетельствует об образе жизни вида.
ПОПУЛЯЦИИ
ПОНЯТИЕ О ПОПУЛЯЦИИ В ЭКОЛОГИИ
Популяцией в экологии называют группу особей одного вида, находящихся во взаимодействии между собой и совместно населяющих общую территорию.
Слово «популяция» происходит от латинского «популюс» — народ, население. Экологическую популяцию, таким образом, можно определить как население одного вида на определенной территории.
Члены одной популяции оказывают друг на друга не меньшее воздействие, чем физические факторы среды или другие обитающие совместно виды организмов. В популяциях проявляются в той или иной степени все формы связей, характерные для межвидовых отношений, но наиболее ярко выражены мутуалистические (взаимно полезные) и конкурентные. Специфические внутривидовые взаимосвязи — это отношения, связанные с воспроизводством: между особями разных полов и между родительским и дочерним поколениями.
При половом размножении обмен генами превращает популяцию в относительно целостную генетическую систему. Если перекрестное оплодотворение отсутствует и преобладает вегетативное, партеногенетическое или другие способы размножения, генетические связи слабее и популяция представляет собой систему клонов, или чистых линий, совместно использующих среду. Такие популяции объединены восновном экологическими связями. Во всех случаях в популяциях действуют законы, позволяющие таким образом использовать ограниченные ресурсы среды, чтобы обеспечить оставление потомства. Достигается это в основном через количественные изменения населения. Популяции многих видов обладают свойствами, позволяющими им регулировать свою численность.
Поддержание оптимальной в данных условиях численности называют гомеостазом популяции. Гомеостатические возможности популяций по-разному выражены у различных видов. Осуществляются они также через взаимоотношения особей.
Таким образом, популяции, как групповые объединения, обладают рядом специфических свойств, которые не присущи каждой отдельно взятой особи. Групповые особенности,— это основные характеристики популяции. К ним относятся: 1) численность — общее количество особей на выделяемой территории;, 2) плотность популяции — среднее число особей на единицу площади или объема занимаемого популяцией пространства; плотность популяции можно выражать также через массу членов популяции в единице пространства; 3) рождаемость — число новых особей, подвившихся за единицу времени4 в результате размножения; (4) смертность— показатель, отражающий количество погибших в популяции особей за определенный отрезок времени; 5) прирост популяции — разница между рождаемостью и смертностью; придет может быть как положительным, так и отрицательным; 6) темп роста — средний прирост за единицу времени.
Популяции свойственна определенная организация. Распределение особей по территории, соотношения групп по полу, возрасту, морфологическим, физиологическим, поведенческим и генетическим особенностям отражают структуру популяции. Она формируется с одной стороны, на основе общих биологических свойств вида, а с другой — под влиянием абиотических факторов среды и популяций других видов. Структура популяций имеет, следовательно, приспособительный характер. Разные популяции одного вида обладают как сходными особенностями структуры, так и отличительными, характеризующими специфику экологических условий в местах их обитания.
Таким образом, кроме адаптивных возможностей отдельных особей, население вида на определенной территории характеризуется еще и приспособительными чертами групповой организации, которые являются свойствами популяции как над индивидуальной системы. Адаптивные возможности вида в целом как системы популяций значительно шире приспособительных особенностей каждой конкретной особи.
ПОПУЛЯЦИОННАЯ СТРУКТУРА ВИДА
Каждый вид, занимая определенную территорию (ареал) представлен на ней системой популяций. Чем сложнее расчленена территория, занимаемая видом, тем больше возможностей для обособления отдельных популяций. Однако в не меньшей степени популяционную структуру вида определяют его биологические особенности, такие, как подвижность составляющих его особей, степень их привязанности к территории, способность преодолевать естественные преграды.
Степень обособленности популяций. Если члены вида постоянно перемещаются и перемешиваются па обширных пространствах, такой вид характеризуется небольшим числом крупных популяций. Большими миграционными способностями отличаются, например, северные олени и песцы. Результаты мечения показывают, что песцы перемещаются за сезон па сотни, а иногда более чем на тысячу километров от мест размножения. Северные олени совершают регулярные сезонные кочевки также в масштабах сотен километров. Границы между популяциями таких видов проходят обычно по крупным географическим преградам: широким рекам, проливам, горным хребтам и т. п. В некоторых случаях подвижный вид при относительно небольшом ареале может быть представлен одной единственной популяцией, например кавказский тур, стада которого постоянно кочуют по двум основным хребтам этого горного массива.
При слабо развитых способностях к перемещению в составе вида формируется множество мелких популяций, отражающих мозаичность ландшафта. У растений и малоподвижных животных число популяций находится в прямой зависимости от степени разнородности среды. Например, в горных районах территориальная дифференцировка таких видов всегда более сложна, чем на ровных открытых пространствах. Примером вида, у которого множественность популяций определяется не столько дифференцировкой среды, сколько особенностями поведения, является бурый медведь. Медведи отличаются большой привязанностью к местам своего обитания, поэтому в пределах обширного ареала представлены множеством относительно мелких группировок, отличающихся друг от друга по ряду свойств.
Степень обособленности соседних популяций вида очень различна. В некоторых случаях они резко разделены территорией, непригодной для обитания, и четко локализованы в пространстве, например популяции окуня и линя в изолированных друг от друга озерах или популяции пластинчатозубой крысы, белоусой славки, индийской камышевки и других видов в оазисах и долинах рек среди пустынь.
Противоположный вариант — сплошное заселение видом обширных территорий. Такой характер распространения свойствен, например, малым сусликам в сухих степях и полупустынях. В этих ландшафтах плотность их населения повсеместно высока. Отдельные непригодные для жизни участки легко преодолеваются при расселении молодняка, а в благоприятные годы на них возникают временные поселения. Здесь вычленить границы между популяциями можно лишь условно, между областями с разной плотностью населения.
Другой пример сплошного распространения вида — семиточечная божья коровка. Эти жуки встречаются в самых различных биотопах и разных природных зонах. Вид характеризуется, кроме того, предзимними миграциями. Границы между популяциями в таких случаях почти не выражены. Однако, поскольку совместно обитающие особи контактируют между собой чаще, чем с представителями других частей ареала, население удаленных друг от друга мест можно считать разными популяциями.
В пределах одного и того же вида могут быть популяции как с хорошо различимыми, так и со смазанными границами. У известного вредителя зерновых культур на юге нашей страны — клопа черепашки — через 2,5—3 месяца после выхода из яиц и активного питания на полях начинается отлет в леса предгорий, за десятки, а порой и сотни километров, где клопы залегают до следующей весны под слой древесного спада. Дальность перелетов зависит от степени упитанности особей, и в результате на одних и тех же зимовках смешиваются клопы,из; разных мест. Направление весеннего разлета сильно зависит от направления ветра, и клопы не обязательно возвращаются в места отрождения. Кроме того, часть черепашек не улетает далеко, а зимует в ближайших лесополосах. Таким образом, на обширных территориях происходит постоянное перемещение и перемешивание особей, которое не позволяет строго разграничивать отдельные популяции. Между тем в определенных частях ареала, как горных, так и равнинных, черепашка ведет относительно оседлый образ жизни, совершая лишь короткие перелеты к ближайшим местам зимовок. В таких районах возникают обособленные популяции, представляющие собой отдельные локальные очаги размножения вредителя. Таким образом, в пределах вида популяции могут быть представлены группами разного объема.
Между популяциями осуществляется обмен отдельными особями, который может быть либо достаточно регулярным, либо эпизодическим. При сезонных кочевках ворон, например, часть молодых птиц ежегодно остается на местах зимовок, образуя пары с представителями оседлого населения. Связь между населением отдельных видов рыб в озерах осуществляется значительно реже, например в годы с особо сильными паводками, когда обособленные водоемы соединяются в единую водную систему.
Взаимоотношения видов не сводятся только к взаимодействию особей, хотя и осуществляются через индивидуальные контакты. Отношения между хищником и его жертвой кратковременны. Связь между их популяциями длительна и является одним из условий функционирования сообществ. Связи между отдельными особями приводят, к одним результатам, связи между популяциями— к качественно другим. Например, длительное воздействие конкретного паразита может иметь следствием изменение физиологического состояния, плодовитости, долговечности хозяина и т. п. Взаимоотношения популяций этих же видов приводят к изменениям их основных групповых характеристик: численности, возрастного состава, смертности, темпов роста популяций.
Связи между популяциями поддерживают вид как единое целое. Слишком длительная и полная изоляция популяций приводит обычно к образованию новых видов.
Различия между отдельными популяциями выражены в разной степени. Они могут затрагивать не только их групповые характеристики, но и качественные особенности физиологии, морфологии и поведения отдельных особей. Эти различия создаются в основном под влиянием естественного отбора, приспосабливающего каждую популяцию к конкретным условиям ее существования.
Зайцы-беляки из разных частей ареала различаются характером окраски, размерами, строением пищеварительной системы. Например, длина слепой кишки у зайцев полуострова Ямал в 2 раза больше, чем у представителей из лесостепного Урала. Это связано с характером питания, разной долей грубых кормов в составе рациона. Средняя величина выводка в центральноякутской популяции составляет около 7 зайчат на самку, которая дает приплод один раз в год. В карельской части ареала зайчихи приносят зайчат дважды за сезон, но средняя величина выводка около 4 экземпляров. В Белоруссии самки беляков могут давать до четырех приплодов в год. Заячье население разных географических территорий испытывает неодинаковое давление со стороны хищников и паразитов. Например, в Верхоянье до 70% зайцев бывает поражено цестодами, вызывающими заболевание мозга. Архангельская популяция больше других страдает от печеночного сосальщика. Различия в характере размножения и масштабах гибели от тех или иных причин определяют различия в общей численности, плотности, темпах роста и структуре популяций разных регионов. Популяции различаются между собой тем сильнее, чем более несходны условия их обитания и чем слабее между ними обмен особями.
Классификация популяций. Экологи руководствуются различными принципами для выделения и классификации популяций как территориальных группировок в пределах вида.
Н. П. Наумов на примере млекопитающих рассматривает вид как иерархическую систему популяций различных рангов. Его классификация основывается на ландшафтно-биотопическом подходе к выделению популяционных единиц. Наиболее крупные территориальные группировки вида — подвиды, или географические расы. Система подвидов и масштабы занимаемой ими территории зависят от биологических особенностей вида. Ареалы подвидов у подвижных форм могут быть очень велики. В их пределах на территориях с однородными географическими условиями выделяются географические популяции, отличающиеся общностью приспособлений к климату и ландшафту. Они, в свою очередь, состоят из более мелких популяций, населяющих различные участки среды. К популяциям низшего ранга применяют разные наименования: экологические, биотопические, местные, локальные, элементарные и т. д. Такие популяции могут быть временными и нестабильными. Чем ниже ранг популяций, тем более тесна связь между соседними популяциями, больше степень обмена особями, менее выражены отличительные особенности. Наиболее сильны различия между группировками подвидового ранга. Они затрагивают не только физиологические и поведенческие черты отдельных особей, но и наследственно закреплены в некоторых чертах их морфологии. Связи между популяциями различных рангов обеспечивают единство вида и обогащение его наследственного фонда.
В школе академика С. С. Шварца развивается другой, историко-генетический подход к выделению природных популяций. С этой точки зрения популяции как генетическое единство можно выделять только у видов с половым размножением и перекрестным оплодотворением. Обязательным признаком популяции считается также ее способность к самостоятельном
|
|
|
