 |


Этологическая структура популяций животных
|
|
|
|
Закономерности поведения животных составляют предмет особой науки — этологии. Систему взаимоотношений между членами одной популяции называют поэтому этологической или поведенческой структурой популяции.
Поведение животных по отношению к другим членам популяции зависит прежде всего от того, одиночный или групповой образ жизни свойствен виду. Формы совместного существования особей в популяции чрезвычайно различны.
Одиночный образ жизни, при котором особи популяции независимы и обособлены друг от друга, характерен для многих видов, но лишь на определенных стадиях жизненного цикла. Полностью одиночное существование организмов в природе не встречается, так как при этом было бы невозможным осуществление их основной жизненной функции — размножения. Однако для некоторых видов характерны очень слабые контакты между совместно живущими особями. Таковы, в частности, отдельные водные обитатели с наружным способом оплодотворения, при котором нет необходимости в непосредственной встрече партнеров, например одиночные актинии. У видов с внутренним оплодотворением встречи самцов и самок также могут быть очень кратковременными, лишь для осуществления копуляции, в, остальное время животные лишь живут независимо друг от друга. Таков образ жизни многих насекомых, например божьей коровки, хищных жуков-жужелиц и др.
У видов с одиночным образом жизни часто образуются временные скопления особей — в местах зимовок, в период, предшествующий размножению, и т. п. Так, бабочки-крапивницы поздней осенью целыми гроздьями собираются в чердачных помещениях или других укрытиях, жужелицы и божьи коровки — в сухой подстилке возле пней и комлей деревьев, сомы и щуки — в зимовальных ямах на дне водоема. У бентосных животных такие скопления могут быть постоянными, например устричные или мидиевые банки, агрегации полихет или поселения морских желудей. В этих поселениях увеличиваются шансы на возможность оплодотворения половыми продуктами, выбрасываемыми в воду. Однако подобные агрегации не сопровождаются установлением более или менее закономерного поведения животных по отношению друг к другу, и каждое из них относительно независимо от остальных.
|
|
|
Дальнейшее усложнение отношений внутри популяции осуществляется по двум направлениям: усилению связи между половыми партнерами и возникновению контактов между родительским и дочерним поколениями.. На этой основе в популяциях формируются семьи, которые очень разнообразны по составу и длительности существования.
Родительские пары могут создаваться как на короткий, так и на длительный срок, а у некоторых видов — на всю жизнь взрослых особей. Например, среди птиц полигамные тетерева, глухари спариваются со многими самками, не образуя устойчивых семейных пар. У ряда уток — кряквы, шилохвости и др.— пары подбираются еще на местах зимовок или во время пролета, но, после того- как самка садится на гнездо, самец ее покидает. У большинства воробьиных самец и самка держатся вместе в течение всего периода гнездования. Семейные пары лебедей, журавлей, голубей сохраняются на долгие годы.
Выбор партнеров сопровождается у животных особым брачным поведением, часто достигающим большой сложности. У пауков-скакунчиков самцы добиваются спаривания с самкой после своеобразного танца, в продолжение которого они описывают возле нее суживающиеся полукруги, принимая особые позы, а приблизившись начинают быстро вертеться, увлекая самку. Считают, что предкопуляционное поведение самцов пауков тормозит инстинкт агрессии у самок, которые иначе реагируют на них как на добычу. Дальнейшее поведение пауков различно: у многих всеядных видов самец после введения им спермы в половые пути самки становится ее жертвой, у бегающих Lycosidae— поспешно спасается бегством, у большинства скакунчиков Salticidae самец и самка расходятся спокойно, а у ряда видов семейства Dictinidaeнекоторое время мирно уживаются в одном гнезде.
|
|
|
Широко известное «ухаживание» у птиц и млекопитающих также предотвращает агрессивные и оборонительные реакции особей противоположного пола, приводит к синхронизации процессов полового созревания и стимулирует у партнеров готовность к спариванию. Это имеет большое значение для осуществления размножения, к которому самец и самка должны быть готовы в одно и то же время.
Выбор половых партнеров часто сопровождается усилением конкурентных отношений в популяциях животных. В период, предшествующий размножению, у многих видов возникают драки самцов, ритуальные демонстрации и другие типы специализированного поведения, направленного на устранение конкурентов. Эти столкновения, несмотря на частую ожесточенность, редко приводят к серьезным ранениям соперников, обычно ограничиваясь изгнанием одного из них с территории, где находятся самки. У рыб, например, в таких сражениях преобладает ритуал угрозы или противники кусают друг друга в пасть — наименее уязвимую часть тела, но никогда не наносят более опасных укусов в бок. Токующие турухтаны стремительно бросаются друг на друга, но затем внезапно замирают с взъерошенным «воротником», после чего вновь повторяют броски.
Таким образом, период, предшествующий размножению в популяциях животных, характеризуется активным поиском и резким усилением контактов между особями.
Семейный образ жизни. При семейном образе жизни усиливаются также связи между родителями и их потомством. Простейший вид такой связи — забота одного из родителей об отложенных яйцах: охрана кладки, инкубация, дополнительное аэрирование и т. п. Так, среди земноводных мраморная амбистома откладывает яйца в ямки во влажной лесной подстилке и остается их сторожить. Самка темной саламандры обвивает кладку своим телом и не покидает ее до выклева личинок. Самец трехиглой колюшки после охраны кладки и заботы о ней некоторое время оберегает и мальков. У птиц сложные заботы о птенцах продолжаются до поднятия их на крыло, а у ряда крупных млекопитающих, например у медведей, тигров, детеныши воспитываются в семейных группах в течение нескольких лет, до наступления их половой зрелости.
|
|
|
Различают семьи отцовского, материнского и смешанного типа, в зависимости от того, кто из родителей берет на себя уход за потомством. В семьях с устойчивым образованием пар в охране и выкармливании молодняка принимают обычно участие и самец и самка.
При семейном образе жизни территориальное поведение животных выражено наиболее ярко: различные сигналы, маркировка, ритуальные формы угрозы и прямая агрессия обеспечивают владение участком, достаточным для выкармливания потомства.
Более крупные объединения животных — стаи, стада и колоний. В основе их формирования лежит дальнейшее усложнение поведенческих связей в популяциях.
Колонии. Это групповые поселения оседлых животных. Они могут существовать длительно или возникать лишь на период размножения, как, например, у многих птиц — грачей, чаек, гагар, тупиков и т. п. По сложности взаимосвязей между особями колонии животных чрезвычайно разнообразны — от простых территориальных скоплений одиночных форм до объединений, где отдельные члены выполняют, как органы в целостном организме, разные функции видовой жизни.
Поселения морских желудей, например, можно рассматривать и как агрегации одиночных особей, и как элементарную форму колонии, так как одновидовые скопления возникают у них не случайно, а на основе хемотаксиса, свойственного личинкам. Взаимоотношения между особями в таких примитивных объединениях сводятся к стимуляции синхронного полового созревания химическими выделениями членов колонии, а конкуренция за облавливаемое пространство в известной мере предотвращается ориентировочной реакцией личинок перед прикреплением.
|
|
|
Более сложная форма колонии — такие поселения животных, в которых некоторые функции их жизни выполняются сообща, что увеличивает вероятность выживания отдельных особей. Чаще всего такими общими функциями становятся защита от врагов и предупредительная сигнализация. Чайки, кайры, некоторые гуси, ласточки и другие птицы обычно с шумом набрасываются на хищника, угрожающего птенцам или кладкам. Тревога, поднятая любой заметившей опасность птицей, мобилизует остальных. Сообща птицам удается изгонять даже крупных хищников, с которыми они не справились бы поодиночке,— песцов, ястребов, сов и др.
В колониальных поселениях птиц часто сохраняются индивидуальные гнездовые участки. У серебристых чаек, например, колонии рыхлые — между гнездами сохраняется расстояние в 3— 5 м. При выборе участков затеваются драки. В. поселениях некоторых других птиц подобные территориальные инстинкты не проявляются совсем. Например, городские ласточки лепят свои гнезда часто вплотную одно к другому.
Общественные ткачики (Рyilacterus socius) строят на деревьях большое общее гнездо из травы с многочисленными отверстиями, ведущими в индивидуальные гнездовые полости. Южноамериканские кукушки-личинкоеды ани строят совместные гнезда, в которые откладывают яйца около десятка самок. Эту общую кладку насиживают одновременно несколько птиц, периодически сменяемые другими.
 Среди млекопитающих колониальны сурки, вискачи, пищухи, пеструшки. Колонии млекопитающих чаще возникают не как территориальные объединения разных семей, а на основе разрастания семейных групп, с сохранением связей между отпочковывающимися семьями. Зверьков из других колоний при полной занятости территории изгоняют, но в период низкой численности популяции они могут вливаться в состав поселения, которое, таким образом, приобретает смешанный характер.
Среди млекопитающих колониальны сурки, вискачи, пищухи, пеструшки. Колонии млекопитающих чаще возникают не как территориальные объединения разных семей, а на основе разрастания семейных групп, с сохранением связей между отпочковывающимися семьями. Зверьков из других колоний при полной занятости территории изгоняют, но в период низкой численности популяции они могут вливаться в состав поселения, которое, таким образом, приобретает смешанный характер.
Наиболее сложные колонии у общественных насекомых —термитов, муравьев, пчел. Они возникают на основе сильно разрастающейся семьи. В таких колониях-семьях насекомые выполняют сообща большинство основных функций: размножения, защиты, обеспечения кормом себя и потомства, строительства и т. п. При этом существует обязательное разделение труда и специализация отдельных особей или возрастных групп на выполнении определенных операций. Члены колонии действуют на основе постоянного обмена информацией друг с другом.
У рыжих лесных муравьев колонии могут занимать одно или несколько гнезд, связанных между собой системой дорог. Самцы выполняют только функцию оплодотворения самок, после чего погибают. Самки сначала осуществляют расселение, затем основывают новую семью, выполняя все операции, связанные с воспитанием первого потомства. После этого они специализируются только на производстве яиц, а остальную работу в муравейнике выполняют рабочие — особая каста, представленная бесплодными измененными самками. Среди них также существует разделение труда. Определенные группы обеспечивают сбор строительного материала, уход за тлями и сбор пади, охоту и разведку, транспортировку добычи, обмен молодью между гнездами и т. п. Кроме того, у рабочих муравьев существует и возрастная смена функций. В течение 1 —1,5 месяца жизни они работают внутри гнезда, ухаживают за молодью, а позднее становятся строителями, разведчиками, фуражирами.
|
|
|
Среди муравьев одной колонии постоянно осуществляется обмен пищей. Муравьи кормят друг друга содержимым зобика, желудка, слизывают выделения различных желез. Это явление получило название трофаллаксиса. На основе трофаллаксиса объединение семьи поддерживается не только общим пищевым потоком, но и действием различных биологически активных веществ, содержащихся в выделениях.
В организации совместной деятельности у муравьев большую роль играет достаточно разнообразная система сигнализации: позами, движениями усиков, прикосновениями разного характера, пахучими выделениями и т. п.
Таким образом, по мере усложнения колониального объединения поведение, а иногда также физиология и строение отдельной особи все больше подчиняются интересам всей колонии.
Особые формы колониальное развиваются у животных с вегетативным размножением, в частности у кишечнополостных и некоторых других водных обитателей. Разделение функций у полипов, связанных между собой системой распределительных каналов, заходит у некоторых видов настолько далеко, что вся колония выглядит и ведет себя как единая особь. Наиболее наглядный пример — сифонофоры, в строении которых практически стираются границы между колонией и цельным организмом, имеющим систему дифференцированных органов. Образование таких колоний-организмов происходит па основе единого метаболизма.

|
Стаи. Это временные объединения животных, которые проявляют биологически полезную организованность действий. Стаи об легчают выполнение каких-либо функций в жизни вида: защиты от врагов, добычи пищи, миграции. Наиболее широко стайность распространена сред птиц и рыб, у млекопитающих характерна для многих собачьих. В стаях сильно развиты подражательные реакции и ориентация на соседей.
По способам координации действий стаи делятся на две категории: I) эквипотенциальные, без выраженного доминирования отдельных членов и 2) стаи с лидерами, в которых животные ориентируются на поведение одной или нескольких, обычно наиболее опытных, особей. Объединения первого типа характерны в основном для рыб, но известны также у мелких птиц, перелетной саранчи и некоторых других форм. Второй тип стай встречается обычно у крупных птиц и млекопитающих.
Стаи рыб очень изменчивы по величине, форме, плотности. Они часто переформировываются, иногда по нескольку раз в сутки. Обычно рыбы группируются в стаи лишь в светлое время суток, при зрительном контакте с другими особями, и рассредоточиваются на ночь. Защитная роль стайных объединений рыб очень велика. В опытах одиночные рыбки вылавливаются хищником в несколько раз быстрее, чем члены стаи. В группе осуществляется «круговой обзор», благодаря которому хищнику труднее приблизиться незамеченным. Кроме того, многочисленность двигающихся особей дезориентирует врага. Тела рыб, непрерывно меняющих направление движения, создают мерцание, затрудняющее фиксацию взгляда на отдельных особях, и делают невозможными целенаправленные броски. Стая быстро маневрирует при опасности, обтекая  хищника, который, бросившись в ее середину, оказывается в пустоте. Для поведения рыб в стае характерен имитационный рефлекс — подражание действиям соседей.
хищника, который, бросившись в ее середину, оказывается в пустоте. Для поведения рыб в стае характерен имитационный рефлекс — подражание действиям соседей.
У птиц стаи формируются при сезонных перелетах или, у оседлых и кочующих форм, при зимних кормежках. Стаи при перелетах образуют те виды, которым свойственно колониальное гнездование или коллективное кормление. Одиночно гнездящиеся и кормящиеся виды стай в полете не образуют.
В стаях оседлых птиц существует постоянная сигнализация, звуковая и зрительная связь между особями. Благодаря внутри-стайной, а часто и межстайной сигнализации птицы используют и опыт, и случайное обнаружение отдельными особями благоприятных мест ночевок и отдыха, источников корма, восходящих потоков воздуха и т. п.
Волчьи стаи возникают для групповой охоты зимой. В группе зверям удается справиться с крупными копытными, охота на которых в одиночку безрезультатна. При групповой охоте волков часто практикуется преследование с выходом на перехват жертвы, нагон жертвы на засаду или захват ее в кольцо, что требует согласованности и координации действий всех особей. Стайность известна для гиен, гиеновых собак, койотов и др. В стаях млекопитающих велика роль вожаков и специфичны отношения между отдельными особями, что сближает эти групповые образования со стадами.
Стада. Это более длительные и постоянные объединения животных по сравнению со стаями. В стадных группах, как правило, осуществляются все основные функции жизни вида: добывание корма, защита от хищников, миграции, размножение, воспитание молодняка и т. п. Основу группового поведения животных в стадах составляют взаимоотношения доминирования-подчинения, основанные на индивидуальных различиях между особями.
Один из вариантов организации стад — это группы с временными либо относительно постоянными лидерами — особями, на которых концентрируется внимание других и которые своим поведением определяют направление перемещения, места кормежки, реакцию на хищников и другие свойства стада. Стадо действует как единое целое, подражая лидеру. Деятельность лидера не направлена непосредственно на подчинение других, особей. Лидером становится более опытный член стада. Например, стада северных оленей обычно ведут старые важенки. Они лучше других ориентируются при миграциях и нападении хищников, так как периодически им приходится делать это в одиночку. Перед рождением олененка самка обычно удаляется и несколько дней, пока не окрепнет малыш, вынуждена одна оберегать и защищать его, а затем вместе с ним догоняет стадо. При переменном лидерстве одни животные становятся во главе стада чаще других. Большое значение имеют при этом также особенности высшей нервной деятельности. В стадах северных оленей около 18—20% животных являются потенциальными лидерами. У других животных лидеры могут быть более постоянны.

|
Биологическое значение лидерства в том, что индивидуальный опыт отдельных особей может быть использован всей группой.
В крупных стадах выделяются также объединения, представляющие семейные или возрастные группы, внутри которых контакты более дружелюбны, чем с членами других аналогичных групп. Во внутристадных группировках, независимо от общего лидерства, могут складываться отношения доминирования-подчинения.
Наиболее сложна поведенческая организация стад с вожаками и иерархическим соподчинением особей. Вожаки, в отличие от лидеров, характеризуются поведением, непосредственно направленным на активное руководство стадом: специальными сигналами, угрозами и прямым нападением. В подобных стадах часто возникают разделение «прав» и «обязанностей» и более сложные формы общественного поведения, выгодные для групп в целом.
Ранг каждой особи в стаде определяется многими причинами. Имеют значение возраст, физическая сила, опыт и наследственные качества животного. Более сильные и опытные, с устойчивым типом нервной системы, как правило, доминируют над более слабыми. Доминирование проявляется в преимуществе при поедании пищи, праве на самку, передвижении в группе и т. п.
Системы доминирования-подчинения очень различны у разных видов. При линейной иерархии в ряду рангов А—В—С и т. д. особи, принадлежащие к каждому рангу, подчинены предыдущим, но главенствуют над последующими. Последние в таком ряду животные— самые бесправные в группе. Подобные отношения возникают, например, в стаях ездовых собак, где вожаки активно подчиняют себе стаю, угрожая и задавая трепку непослушным. Животные низшего ранга проявляют признаки покорности перед всеми остальными, подходят к пище в последнюю очередь, их не подпускают к самкам, изгоняют с лучших мест отдыха и т. д. Иерархия в группах часто бывает более сложной. Например, могут возникать параллельные ряды подчинения: один среди самцов, другой — среди самок, как, например, у некоторых игрунковых обезьян. У павлинов, помимо того, самцы доминируют над самками, а те в свою очередь над детенышами. У некоторых видов обнаружено иерархическое соподчинение по типу «треугольника»: А нападает на В, В — на С, а С подчиняет себе А. Такое, соотношение может сохраняться в группе довольно долго. Одним из вариантов иерархии может быть деспотия — доминирование одного животного над всеми остальными членами группы.
Ранг животного в группе определяется столкновениями между особями, которые могут иметь характер прямой борьбы или ритуальных угроз. После установления ранга всех членов группы прямые столкновения между ними прекращаются и порядок поддерживается сигнальным или ритуальным поведением.
Иерархически организованному стаду свойствен закономерный порядок перемещения, определенная организация при защите, расположении на местах отдыха и т. п. Так, при передвижении семейного стада зебр впереди всегда идет старая кобыла, за нею следуют сначала самые молодые, а затем старшие жеребята, за которыми двигаются взрослые зебры. Шествие завершает жеребец-вожак. В стаде павианов в центре, в наибольшей безопасности, находятся самки с детенышами или готовящиеся к размножению, по краям — вожаки, молодые самцы и неразмножающиеся самки.

Впереди и позади стада шествуют крупные самцы, готовые отравить нападение. Описаны случаи, когда при преследовании стада хищниками самец-доминант возвращался за отставшим детенышем несмотря на грозившую ему большую опасность.
Вожаки-доминанты выполняют разнообразные общественные функции. В табуне лошадей, например, вожак управляет движением, уводит косяк от опасности, защищает от хищников, прекращает драки, проявляет заботу о жеребятах и больных животные и тп п. В сложной обстановке вожаки ряда видов (лошади Пржевальского, павианы) могут проводить разведку, отделяясь от стада, а затем возвращаясь к нему и определяя безопасное направление движения.
Конфликты в группах, имеющих вожака, обычно решаются с его участием или в его присутствии, у разных видов с разной степенью агрессивности. В стадах павианов доминанты могут прибегать к преследованию и укусам, тогда как у горилл вожак нередко восстанавливает порядок лишь взглядом или движением головы.
Биологический смысл иерархической системы доминирования-подчинения заключается в создании согласованного поведения группы, выгодного для всех ее членов. После «расстановки сил» животные не тратят лишней энергии на индивидуальные конфликты, а группа в целом получает преимущества, подчиняясь наиболее сильным и опытным индивидуумам. Согласованность поведения имеет большое значение в обеспечении защиты от хищников, предупреждении опасности, выращивании молодняка, миграциях и т. п. В сложных условиях, например при голодовках, гибнут в первую очередь более слабые, подчиненные особи, но под защитой группы они все же имеют больше шансов выжить, чем в одиночку.
Иерархия в групповых отношениях адаптивна прежде всего потому, что она динамична. В жизни стада идет непрерывная проверка соответствия его структуры меняющимся условиям. Сбегав группы также не остается постоянным: взрослеет молодняк, появляются новые пришельцы, старые животные теряют силы, часть особей погибает. Эти изменения сопровождаются перестройкой рангов. Переходу особи в новый ранг всегда предшествуют прямые или ритуальные схватки и демонстрации, «пересмотр сил», после чего закрепляется новое положение. На роль вожака-доминанта всегда претендуют более молодые, набирающие силу и опыт животные. Они занимают его место после первого же проявления вожаком неспособности отстоять свои права.
Иерархия наиболее ярко выражена в стадах, но она характерна и для многих других групп оседлых животных — семей, колоний. Иерархия может проявляться также в распределении территориальных участков у одиночных форм. Среди сусликов, охраняющих свои индивидуальные территории, доминирующие особи занимают наиболее благоприятные места в поселениях. Среди колониальных птиц (чаек, гусей, цапель и др.) наиболее крупные и опытные гнездятся в центре колонии, тогда как периферию занимают физиологически менее полноценные особи, проявляющие реакцию подчинения по отношению к доминантам.

|
Система доминирования-подчинения выявлена и у ряда беспозвоночных: насекомых (сверчков, жуков-чернотелок и др.), некоторых ракообразных и пр.
Установление иерархии сопровождается изменениями в гормональной системе организмов. У млекопитающих, например, возникает гипертрофия коры надпочечников. Эти изменения оказываются наибольшими у животных, находящихся в самом низу иерархической лестницы, и не проявляются у домипантов. Стероидные гормоны, выделяемые корой надпочечников, оказывают большое влияние на процессы обмена веществ, которые, в свою очередь, лежат в основе изменения поведения.
Эффект группы. Жизнь в группе через нервную и гормональную системы отражается на протекании многих физиологических процессов в организме животного. У изолированных особей заметно меняется уровень метаболизма, быстрее тратятся резервные вещества, не проявляется целый ряд инстинктов и ухудшается общая жизнеспособность.
Оптимизация физиологических процессов, ведущая к повышению жизнеспособности при совместном существовании, получила название «эффект группы». Он проявляется как психофизиологическая реакция отдельной особи на присутствие других особей своего вида. У овец вне стада учащаются пульс и дыхание, а при виде приближающегося стада эти процессы нормализуются. Одиночно зимующие летучие мыши отличаются более высоким уровнем обмена веществ, чем в колонии. Это ведет к повышенной трате энергии, истощению и часто заканчивается гибелью животных.

|
Затраты организмом энергии на основную жизнедеятельность учитывают, измеряя скорость потребления кислорода животными в покое. Эти показатели возрастают у всех животных, приспособленных к групповой жизни и оказавшихся вне стада, семьи или колонии Напротив, даже простое скучивание сокращаут газообмен порой до 50%. Возможно, что это явление лежит в основе агрегации на зимовку рыб, лягушек, моллюсков, насекомых, у которых также обнаружена экономия энергетических затрат в скоплениях.
Эффект группы проявляется в ускорении темпов роста животных, повышении плодовитости, более быстром образовании условных рефлексов, повышении средней продолжительности жизни индивидуума и т. д. В группе животные часто способны поддерживать оптимальную температуру (при скучивании, в гнездах, в ульях). У многих животных вне группы не реализуется-плодовитость. Голуби некоторых пород не откладывают яйца, если не видят других птиц. Достаточно поставить перед самкой зеркало, чтобы она приступила к яйцекладке. Кайры при небольшой численности пар в колонии начинают размножение только тогда, когда колония увеличится по периферии за счет других видов: чистиков, тупиков,
моевок.
Эффект группы не проявляется у видов, ведущих одиночный образ жизни. Если таких животных искусственно заставить жить (вместе, у них повышается раздражительность, учащаются столкновения и многие физиологические показатели сильно уклоняются от оптимума, о чем можно судить, например, по увеличению энергетических затрат на основные процессы жизнедеятельности. Так, ушастые ежи в группе повышают потребление кислорода до 134% по сравнению с содержащимися поодиночке.
Положительный эффект группы проявляется лишь до некото- -рого оптимального уровня плотности популяции. Если животных становится слишком много, это грозит для всех недостатком ресурсов среды. Тогда вступают в действие другие механизмы, приводящие к снижению численности особей в группе путем ее деления, рассредоточения или падения рождаемости. Эти механизмы обсуждены ниже, при рассмотрении гомеостаза популяций.
Динамика популяций
Биотический потенциал. Любая популяция теоретически способна к неограниченному росту численности, если ее не лимитируют факторы внешней среды. В таком гипотетическом случае скорость роста популяции будет зависеть только от величины биотического потенциала, свойственного виду. Понятие биотического потенциала введено в экологию в 1928г. Р. Чепменом. Этот показатель отражает теоретический максимум потомков от одной пары (или одной особи) за единицу времени, например за год или за весь жизненный цикл.
При расчетах его чаще всего выражают коэффициентом r и вычисляют как максимально возможный прирост популяции Nза отрезок времени t, отнесенный к одной особи, при начальной численности популяции N:
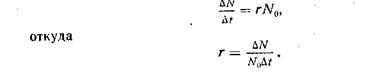
|
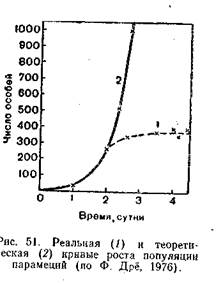
Величина биотического потенциала чрезвычайно различна у разных видов. Например, самка косули способна произвести за жизнь 10—15 козлят, трихина — отложить 1,8 тыс. личинок, самка медоносной пчелы — 50 тыс. яиц, а луна-рыба до 3 млрд. икринок. Биотический потенциал этих видов на самом деле еще больше, так как часть развивающихся яиц и зародышей погибает еще до рождения. Если бы все зародыши сохранялись, а все потомство выживало, численность любой популяции через определенные интервалы увеличивалась бы в геометрической прогрессии.
Кривая, отражающая на графике подобный рост популяций, быстро увеличивает крутизну и уходит в бесконечность. Такая кривая носит название экспоненциальной. На логарифмической шкале подобная зависимость численности популяции от времени будет представлена прямой, а биотический потенциал г отразился ее наклоном по отношению к горизонтальной оси, который тем круче, чем больше величина г.
В природе биотический потенциал популяции никогда не реализуется полностью. Его величина обычно складывается как разность между рождаемостью и смертностью в популяциях; r = b-d, где b — число родившихся, а d — число погибших особей в популяции за один и тот же период времени.
|
Общие изменения численности популяции складываются за счет четырех явлений: рождаемости, смертности, вселения и выселения особей (миграция).
Рождаемость. Это число новых особей, появляющихся в популяции за единицу времени в расчете на определенное число ее членов.
Различают абсолютную и удельную рождаемость. Первая характеризуется общим числом родившихся особей. Например, если в популяции северных оленей, насчитывающей 16 тыс. голов, за год появилось 2 тыс. оленят, то это число и выражает абсолютную рождаемость. Удельную вычисляют как среднее изменение численности на особь за определенный интервал времени, и в данном случае она составит 0,125, т. е. один новорожденный на 8 членов популяции за год.
Величина рождаемости зависит от многих причин. Большое значение имеет доля особей, способных в данный период к размножению, что определяется соотношением полов и возрастных групп. Важна также частота последовательности генераций. Так, среди насекомых различают моновольтинные и поливольтинные виды. Первые дают одну, вторые —несколько генераций за год. Например, у тлей за сезон насчитывается до 35 партеногенетических поколений. По числу периодов размножения в течение жизни различают моноциклические и полициклические виды. Моноцикличность свойственна обычно видам с короткой продолжительностью жизни в половозрелом состоянии (лососевые рыбы, поденки, майские жуки и многие другие насекомые). Полицикличность характеризуется повторным размножением особей и присуща большинству позвоночных животных и ряду беспозвоночных, например ракообразным.
У растений выделяют монокарпические и поликарпические виды, т. е. с однократным и многократным размножением в течение жизни. Для величины рождаемости имеет значение также соотношение периода размножения и общей продолжительности жизни. Этот период для самок дрозофил составляет около 65%, для стадной саранчи шистоцерки—15%, а для поденок — всего от 0,5 до 1% длительности их существования. Большую роль играет плодовитость особей. Однако размножаемость популяции, как правило, не бывает прямо пропорциональна плодовитости. Плодовитость сильно зависит от степени развития заботы о потомстве или обеспеченности яиц питательными материалами. Среди рыб наибольшее количество икринок выметывают виды с пелагической икрой— сельди, тресковые, камбаловые и др. Например, сахалинекая сельдь продуцирует 38—46 тыс. мелких, в доли миллиметра, икринок. У лососевых, зарывающих икру в грунт, развивается меньшее число яиц, но более крупных размеров. Средняя плодовитость амурской горбуши 1300—1500 икринок диаметром 4—6 мм. Наиболее крупная икра у акул и химер, до 6—8 см. Яйца этих рыб имеют к тому же плотную защитную оболочку. Количество их Очень невелико — несколько штук на одну самку.
У видов, оберегающих и выкармливающих, свой молодняк, плодовитость резко понижена. Размер кладки у птиц разных видов варьирует уже не в тысячи раз, как у рыб, а в пределах от одного яйца (у некоторых хищников, пингвинов, чистиков и др.) до 20—25 (максимальное количество яиц у куриных, например, у серой куропатки).
Д. Л э к установил на примере птиц, что у них отбор благоприятствует не максимальной плодовитости, а наиболее эффективной, т. е. тому количеству яиц, при котором потомство оказывается наиболее жизнеспособным. На выкармливание птенцов птицы тратят огромное количество энергии. Мелкие птицы приносят пищу в гнездо сотни раз в сутки: горихвостка — более 200, большая синица — около 400, а крапивник — до 600 раз. Если величина кладки больше обычного предела, птенцы оказываются недокормленными и их жизнеспособность снижена.

|
Большая плодовитость вырабатывается у видов также в условиях более высокой смертности, особенно при сильном давлении со стороны хищников. Отбор на более высокую плодовитость — одно из приспособлений, компенсирующих большую норму гибели в популяциях.
Поэтому при высокой плодовитости общая размножаемость популяции может быть весьма низкой. В различиных популяциях одного вида плодовитость обычно тем выше, чем более неблагоприятны занимаемые ими места обитания. Так, у многих млекопитающих — зайцев, мышей, полевок — число детенышей в помете па границах ареала больше, чем в его центре.
Смертность. Смертность в популяциях также зависит от многих - причин: генетической и физиологической полноценности особей, влияния неблагоприятных физических условий среды, воздействия хищников, паразитов, болезней и т. п. На разных стадиях жизненного цикла каждого поколения эти факторы действуют с разной силой. На практике, когда необходимо проанализировать ход смертности в популяциях, составляют таблицы выживания (см. далее), где для каждой возрастной группы указываются эмпирически полученные данные, характеризующие гибель особей данного возраста. На основе таких таблиц составляют кривые выживания, позволяющие прогнозировать в сходных условиях состояние очередных генераций. {

|
Идеальный случай, если все особи одной генерации доживают до биологически предельного возраста, а затем в течение короткого срока отмирают. Этому соответствует минимальная смертность популяции.
Кривая, описывающая такую зависимость численности поколений от времени, сначала идет параллельно горизонтальной оси, а затем круто загибается вниз. В природе подобный ход смертности не свойствен ни одному виду, но встречается приближение к нему, например, у таких насекомых с хорошо защищенными личинками, как жуки-златки, у крупных млекопитающих с высокой степенью выживаемости молодняка. К этому же типу близка кривая, описывающая смертность в современных популяциях человека. Средняя продолжительность жизни особи в таких популяциях высока и приближается к максимальной.
Найболее часто встречающийся в природе вариант—'Повышенная гибель особей в ранний период жизни. Взрослые формы более защищены или выносливы. Кривая смертности в таких популяциях резко падает к горизонтальной оси в самом начале. Ее наклон отражает скорость убыли поколения. Так, у большинства рыб значительная часть популяции гибнет еще на стадии икры, очень велик также отсев мальков и лишь среди взрослых
|
|
|
