 |


Промбриональный и эмбриональный
|
|
|
|
Периоды
Проэмбриональный (от греч. pro — до, embryon — зародыш) период в индивидуальном развитии организмов связан с образованием половых клеток в процессе гаметогенеза. Как отмечено выше, мужские половые клетки животных по своей структуре не имеют существенных отличий от других (соматических) клеток, тогда как яйцеклетки характеризуются важной отличительной чертой, заключающейся в том, что они содержат очень много желтка. Учитывая количество желтка и топографию его в яйцеклетках, последние классифицируют на три типа, а именно:
1. Изолецитальные клетки. Эти яйцеклетки содержат немного желтка, который локализован равномерно по всей клетке. Изолецитальные яйцеклетки продуцируются иглокожими (морскими ежами), низшими хордовыми (ланцетниками), млекопитающими.
2. Телолецитальные яйцеклетки. Эти яйцеклетки содержат большое количество желтка, который сосредоточен на одном из полюсов — вегетативном. Такие яйцеклетки продуцируются моллюсками, земноводными, рептилиями, птицами. Например, яйцеклетки лягушки состоят из желтка на 50%, яйцеклетки кур (в обиходе куриные яйца) — на 95%. На другом полюсе (анимальном) телоле-ЦИтальных яйцеклеток сосредоточены цитоплазма и ядро.
3. Центролецитальные яйцеклетки. В этих яйцеклетках желтка немного, но он занимает центральное положение. Периферию таких яйцеклеток занимает цитоплазма. Примером центролециталь-ных яйцеклеток являются яйцеклетки, продуцируемые членистоногими.
Для проэмбрионального периода характерно также то, что в этот период в гаметах происходят метаболические процессы, связанные с накоплением интенсивно синтезируемых молекул РНК.
Эмбриональный период или эмбриогенез (от греч. embryon — за-родыш, genesis — развитие), начинается со слияния ядер мужской и Женской половых клеток, который представляет собой процесс оплодотворения яйцеклеток. У организмов, для которых характерно внутриутробное развитие, эмбриональный период заканчивается рождением потомства, а у организмов, для которых характерны личиночный и неличиночный типы развития, эмбриональный период завершается выходом потомства из яйцевых или зародышевых оболочек соответственно.
|
|
|
В пределах эмбрионального периода различают стадии зиготы, дробления, бластулы, гаструлы, образования зародышевых листков, гистогенез и органогенез. Как отмечено выше, с учетом фактора времени у млекопитающих и человека зародыш до момента формирования зачатков органов называют эмбрионом, а после этого вплоть до рождения называют плодом. У человека развитие эмбриона (зародыша) заканчивается к концу второго месяца. Начиная с 9-й недели, следует плодный период, характеризующийся дальнейшим ростом и развитием организма (плода) во внутриутробном состоянии вплоть до рождения.
Зигота. У млекопитающих зигота образуется в результате оплодотворения, начинающегося с того, что одна из мужских половых клеток достигает яйцеклетки и инициирует ее развитие. В активированной мужской половой клеткой яйцеклетке происходит ряд физических и химических процессов, включая перемещение протоплазмы, что ведет к установлению билатеральной симметрии яйцеклетки, а также перестройку плазматической мембраны, что исключает слияние с яйцеклеткой других (дополнительных) мужских половых клеток. Затем следует слияние плазматических мембран яйцеклетки и спермия с последующим разрушением ядерных мембран, что обеспечивает слияние ядер двух клеток. Ядра клеток сливаются, при этом восстанавливается диплоидный набор хромосом. Оплодотворение яйцеклетки сопровождается активированием в ней синтеза белков. Таким образом, образуется по существу одноклеточный организм.
|
|
|
Дробление. Образование морулы. Дробление представляет собой начальный период развития зиготы (оплодотворенного яйца). Поскольку яйцеклетки обладают центриолями, то оно заключается в делении зиготы путем митоза, которое начинается, например у человека, через 30 часов после осеменения. У человека деление начинается с движения оплодотворенной яйцеклетки по фаллопиевой трубе и заключается в появлении на поверхности яйцеклетки борозды. Первая борозда приводит к образованию двух клеток — двух бластомеров, вторая — четырех бластомеров, третья — восьми бластомеров и т. д. (рис. 88). Группа клеток, образовавшаяся в результате последовательных дроблений зиготы, получила название морулы (от греч. morum — тутовая ягода).
Стадию морулы проходят все многоклеточные животные, размножающиеся половым путем. В зависимости от видовой принадлежности деление идет по-разному. Различают радиальное (позвоночные, иглокожие), билатеральное (грибневики, некоторые хордовые) и спиральное дробление (немертины, кольчатые черви, многие моллюски), причем эти формы дробления зависят от плоскостей дробления. Поэтому их морулы состоят из разного количества клеток. Кроме того, из части клеток образуется структура, называемая трофобластом, клетки которого питают зародыш, а благодаря ферментам обеспечивают также внедрение последнего в стенку матки. У человека прикрепление морулы к стенке матки происходит на 7-й день после оплодотворения. Позднее клетки трофобласта отслаиваются от зародыша и образуют пузырек, который заполняется жидкостью тканей матки.
Характерная особенность дробления заключается в том, что при нем значительного роста клеток не происходит. Поэтому биологическое значение этой стадии заключается в том, что из крупной клетки, которой является яйцеклетка, образуются более мелкие клетки, в которых уменьшено отношение цитоплазмы к ядру. В результате этого происходит изменение топологии цитоплазматических комплексов в бластомерах, что создает новое цитоплазматическое окружение для ядер.
Дробление зиготы завершается образованием многоклеточной структуры, получившей название бластулы (от греч. blastos — росток). Эта структура имеет форму пузырька, состоящего из одного слоя клеток, называемого бластодермой. Теперь эти клетки называют эмбриональными. По размерам бластула сходна с яйцеклеткой. В период дробления увеличивается количество ядер, возрастает общее количество ДНК. В конце стадии бластулы синтезируется также небольшое количество мРНК и тРНК, но новые рибосомы и рибосомная РНК до начала гаструляции еще не обнаруживаются, либо если обнаруживаются, то в ничтожных количествах.
|
|
|
Гаструляция. Гаструляция (от греч. gastre — полость сосуда) — это следующий за образованием бластулы процесс движения эмбриональных клеток, который сопровождается формированием двух или трех (в зависимости от вида животных) слоев зародыша или так называемых зародышевых листков (рис. 89).
Гаструляция характеризуется увеличением интенсивности обмена по сравнению с дроблением в 2-3 раза. Резко возрастает синтез мРНК, рРНК, рибосом и белков.
Развитие (гаструляция) изолецитальных яиц происходит путем инвагинации (впячивания) вегетативного полюса внутрь бластулы, в результате чего противоположные полюса почти сливаются, а бла-стоцель (полость бластулы) почти исчезает либо полностью исчезает. Внешний слой клеток зародыша получил название эктодермы (от греч. ectos — снаружи, derma — кожа) или наружного зародышевого листка, тогда как внутренний — энтодермы (от греч. entos — внутри) или внутреннего зародышевого листка. Образующаяся при этом полость получила название гастроцели, или первичной кишки, вход в которую называют бластопором (первичным ртом).


Развитие двух зародышевых листков характерно для губок и кишечнополостных. Однако для хордовых в период гаструляции характерно развитие третьего зародышевого листка — мезодермы (от греч. mesos — средний), образующегося между эктодермой и энтодермой (рис. 90).
Гаструляция является необходимым пререквизитом для последующих стадий развития, поскольку она приводит клетки в положение, открывающее возможность формировать органы. Дифференцированный на три эмбриональных закладки зародышевый материал дает начало всем тканям и органам развивающегося зародыша.
|
|
|
Гистогенез и органогенез
Развитие (дифференцировка) зародышевых листков в ходе эмбриогенеза сопровождается тем, что из них формируются различные ткани и органы. В частности, из эктодермы развиваются эпидермис кожи, ногти и волосы, сальные и потовые железы, нервная система (головной мозг, спинной мозг, ганглии, нервы), рецептор-ные клетки органов чувств, хрусталик глаза, эпителий рта, носовой полости и анального отверстия, зубная эмаль. Из энтодермы развиваются эпителий пищевода, желудка, кишек, желчного пузыря, трахеи, бронхов, легких, мочеиспускательного канала, а также печень, поджелудочная железа, щитовидная, паращитовидная и зобная железы. Из мезодермы развиваются гладкая мускулатура, скелетные и сердечные мышцы, дерма, соединительная ткань, кости и хрящи, дентин зубов, кровь и кровеносные сосуды, брыжейка, почки, семенники и яичники. У человека первыми обособляются головной и спинной мозг. Через 26 дней после овуляции длина человеческого зародыша составляет около 3,5 мм. При этом уже видны зачатки рук, но зачатки ног только вступают в развитие. Через 30 дней после овуляции длина зародыша равна уже 7,5 мм. В это время уже можно различить сегментацию зачатков конечностей, глазные бокалы, полушария головного мозга, печень, Желчный пузырь и даже разделение сердца на камеры.
У восьминедельного зародыша человека при длине его около 40 мм и весе около 5 г появляются почти все структуры тела. Органогенез заканчивается к концу эмбрионального периода. В это время эмбрион по внешнему виду приобретает черты сходства с человеком.
Длина 12 недельного человеческого плода составляет уже около 87 мм, а масса около 45 г. Продолжается дальнейший рост и развитие плода. Например, на 4-м месяце развития появляются волосы, а на 20-й неделе начинают образовываться клетки крови.
Если дефинитивное ротовое отверстие образуется на месте первичного рта (бластопора), то этих животных называют первичноро-тыми (черви, моллюски, членистоногие). Если же дефинитивный рот образуется в противоположном месте, то этих животных называют вторичноротыми (иглокожие, хордовые).
Для обеспечения связи зародыша со средой у него развиваются так называемые провизорные органы, которые существуют временно. В зависимости от типа яйцеклеток провизорными органами являются разные структуры. У рыб, рептилий и птиц провизорным органом является желточный мешок. У млекопитающих желточный мешок закладывается в начале эмбриогенеза, но не развивается. Позднее он редуцируется. В ходе эволюции у рептилий, птиц и млекопитающих развились зародышевые оболочки, обеспечивающие защиту и питание эмбрионов (рис. 91). У млекопитающих, в т. ч. у человека, эти зародышевые оболочки являются листками ткани, развивающимися из тела эмбриона. Существуют три такие оболочки — амнион, хорион и аллантоис. Наружная оболочка эмбриона называется хорионом. Она врастает в матку. Место наибольшего врастания в матку называется плацентой. Зародыш с плацентой связан через пуповину или пупочный канатик, в котором имеются кровеносные сосуды, обеспечивающие плацентарное кровообращение. Амнион развивается из внутреннего листка, а аллантоис развивается между амнионом и хорионом. В пространстве между эмбрионом и амнионом, которое называется амниоти-ческой полостью, содержится жидкость (амниотическая). В этой жидкости находится эмбрион, а затем и плод до самого рождения. Метаболизм плода обеспечивается через плаценту.
|
|
|
В основе формообразующего взаимодействия частей эмбриона лежат определенным образом скоординированные процессы обмена веществ. Закономерностью развития является гетерохронность, под которой понимают разное во времени образование закладок органов и разную интенсивность их развития. Быстрее развиваются те органы и системы, которые должны раньше начать функционировать. Например, у человека зачатки верхних конечностей развиваются быстрее, чем зачатки нижних.

Большое влияние на развитие зародыша и плода оказывают условия жизни матери. Зародыш чрезвычайно чувствителен к разным воздействиям. Поэтому различают так называемые критические периоды, т. е. периоды, в которых зародыши, а потом и плоды наиболее чувствительны к повреждающим факторам. В случае человека критическими периодами эмбрионального онтогенеза являются первые дни после оплодотворения, время образования плаценты и роды, а повреждающими факторами являются алкоголь, токсические вещества, недостаток кислорода, вирусы, бактерии, патогенные простейшие, гельминты и другие факторы. Эти факторы обладают терратогенным действием и ведут к уродствам, нарушениям нормального развития.
 Еще со времен Гиппократа (V в. д. н. э.) обсуждается вопрос о причинах, которые инициируют рождение плода. В частности, сам Гиппократ предполагал, что развитие плода инициирует собственное рождение. Новейшие экспериментальные работы английских исследователей, выполненные на овцах, показали, что у овец инициация окотов контролируется комплексом гипоталамус + гипофиз + надпочечники плодов. Повреждения ядер гипоталамуса, удаление передней доли гипофиза или надпочечников пролонгирует беременности овец. Напротив, введение овцам аденокортикотроп-ного гормона (секрета передней доли гипофиза) или кортизола (секрета надпочечников) сокращает сроки их беременностей.
Еще со времен Гиппократа (V в. д. н. э.) обсуждается вопрос о причинах, которые инициируют рождение плода. В частности, сам Гиппократ предполагал, что развитие плода инициирует собственное рождение. Новейшие экспериментальные работы английских исследователей, выполненные на овцах, показали, что у овец инициация окотов контролируется комплексом гипоталамус + гипофиз + надпочечники плодов. Повреждения ядер гипоталамуса, удаление передней доли гипофиза или надпочечников пролонгирует беременности овец. Напротив, введение овцам аденокортикотроп-ного гормона (секрета передней доли гипофиза) или кортизола (секрета надпочечников) сокращает сроки их беременностей.
Довольно частым нарушением развития является разделение зародыша на очень ранней стадии развития, что сопровождается развитием однояйцевых (моно-зиготных) близнецов (рис. 92). Известны также и так называемые сиамские близнецы, представляющие собой неразделенные организмы. Неразделенность встречается разной — от незначительного соединения до почти полного слияния двух организмов с разделенными головами или ногами. Иногда из двух сиамских близнецов один нормален, но другой чрезвычайно изменен, будучи прикрепленным к нормальному, являясь, по существу, паразитом.
 Итак, в процессе развития высших эукариотов одиночная оплодотворенная клетка-зигота в ходе дальнейшего развития в результате митоза дает начало клеткам разных типов — эпителиальным, нервным, костным, клеткам крови и другим, которые характеризуются разнообразием морфологии и макромолекулярного состава. Однако для клеток разных типов характерно и то, что они содержат одинаковые наборы генов, но являются высокоспециализированными, выполняя лишь одну или несколько специфических функций, т. е. одни гены «работают» в клетках, другие неактивны. Например, только эритроциты специфичны в синтезе и хранении гемоглобина.
Итак, в процессе развития высших эукариотов одиночная оплодотворенная клетка-зигота в ходе дальнейшего развития в результате митоза дает начало клеткам разных типов — эпителиальным, нервным, костным, клеткам крови и другим, которые характеризуются разнообразием морфологии и макромолекулярного состава. Однако для клеток разных типов характерно и то, что они содержат одинаковые наборы генов, но являются высокоспециализированными, выполняя лишь одну или несколько специфических функций, т. е. одни гены «работают» в клетках, другие неактивны. Например, только эритроциты специфичны в синтезе и хранении гемоглобина.
Точно так лишь клетки эпидермиса синтезируют кератин. Поэтому давно возникли вопросы о генетической идентичности ядер соматических клеток и о контрольных механизмах развития оплодотворенных яйцеклеток как пререквизита в познании механизмов, лежащих в основе дифференцировки клеток.
Начиная с 50-х годов во многих лабораториях были выполнены эксперименты по успешной пересадке ядер соматических клеток в яйцеклетки, искусственно лишенные собственных ядер. Исследование ДНК из ядер разных дифференцированных клеток показало, что почти во всех случаях геномы содержат одинаковые наборы последовательностей нуклеотидных пар. Известны лишь исключения, когда эритроциты млекопитающих теряют свои ядра в течение последней стадии дифференцировки. Но к этому времени пулы стойких гемоглобиновых мРНК уже синтезированы, так что ядра больше не нужны эритроцитам. Другими примерами служат гены иммуноглобулинов и Т-клеток, модифицируемые в ходе развития.
Одним из крупных этапов в направлении познания контрольных механизмов эмбрионального онтогенеза стали результаты экспериментов, выполненных в 1960—70 гг. английским исследователем Д. Гёрдоном с целью выяснить, обладают ли ядра соматических клеток способностью обеспечивать дальнейшее развитие яйцеклеток, если эти ядра ввести в яйцеклетки, из которых предварительно удалены собственные ядра. На рис. 93 приведена схема одного из этих экспериментов, в котором ядра соматических клеток головастика пересаживали в яйцеклетки лягушки с предварительно удаленными ядрами. Эти эксперименты показали, что ядра соматических клеток действительно способны обеспечивать дальнейшее развитие яйцеклеток, т. к. они оказались способными оплодотворять яйцеклетки и «заставляли» их развиваться дальше. Этим была показана возможность клонирования животных.
Позднее другими исследователями были выполнены эксперименты, в которых было показано, что перенос отдельных бластомеров из 8- и 16-дневных эмбрионов овец одной породы в безъядерную половину яйцеклетки (после рассечения последней пополам) другой породы сопровождалось формированием жизнеспособных эмбрионов с последующим рождением ягнят.
В начале 1997 г. английскими авторами было показано, что введение в искусственно лишенные ядра яйцеклеток овец ядер соматических клеток (клеток эмбрионов, плодов и вымени взрослых овец), а затем имплантация оплодотворенных таким образом яйцеклеток в матку овец сопровождается возникновением беременности с последующим рождением ягнят.
Оценка этих результатов показывает, что млекопитающих можно размножать асексуальным путем, получая потомство животных, клетки которых содержат ядерный материал отцовского или материнского происхождения в зависимости от пола овцы-донора, в таких клетках лишь цитоплазма и митохондрии имеют материнское происхождение. Это заключение имеет чрезвычайно важное общебиологическое значение, расширяет наши взгляды на потенциал размножения животных. Но важно также добавить, что речь идет о генетических манипуляциях, которые в природе отсутствуют. С другой стороны, в практическом плане эти генетические манипуляции представляют собой прямой путь клонирования высокоорганизованных животных с заданными свойствами, что имеет важное экономическое значение (см. раздел VI). В медицинском плане этот путь, возможно, будет использован в будущем для преодоления мужского бесплодия.
Итак, генетическая информация, необходимая для нормального развития эмбриона, не теряется в течение дифференцировки клеток. Другими словами, соматические клетки обладают свойством, получившим название тотипотентности, т. е. в их геноме содержится вся информация, полученная ими от оплодотворенной яйцеклетки, давшей им начало в результате дифференциации. Наличие этих данных с несомненностью означает, что дифференциация клеток подвержена генетическому контролю.
Установлено, что интенсивный белковый синтез, следующий за оплодотворением, у большинства эукариотов не сопровождается синтезом мРНК. Изучение овогенеза у позвоночных, в частности У амфибий, показало, что интенсивная транскрипция происходит еще в течение профазы I (особенно диплотены) мейоза. Поэтому генные транскрипты в форме молекул мРНК или про-мРНК сохраняются в яйцеклетках в бездействующем состоянии. Установлено, что у эмбриональных клеток имеет место так называемое ассимет-ричное деление, заключающееся в том, что деление эмбриональной клетки дает начало двум клеткам, из которых лишь одна наследует белки, принимающие участие в транскрипции. Таким образом, неравное распределение транскрипционных факторов между дочерними клетками ведет к экспрессии в них разных наборов генов после деления, т. е. к дифференциации клеток.
У амфибий и, возможно, у большинства позвоночных, генетические программы, контролирующие раннее развитие (до стадии бластулы), устанавливается еще в течение овогенеза. Более поздние стадии развития, когда начинается клеточная дифференциация (примерно со стадии гаструлы) нуждаются в новых программах для экспрессии генов. Таким образом, дифференцировка клеток связана с перепрограммированием генетической информации в том или ином направлении.
Характерная особенность дифференцировки клеток заключается в том, что она необратимо ведет к тому или иному типу клеток. Этот процесс носит название детерминации и также находится под генетическим контролем, а как сейчас предполагают, дифференциация и детерминация клеток регулируется взаимодействием клеток на основе сигналов, осуществляемых пептидными ростовыми факторами через тирозинкиназные рецепторы. Вероятно, существует много таких систем. Одна из них заключается в том, что дифференциация мышечных и нервных клеток регулируется нейрорегу-линами, представляющими собой мембранные белки, действующие через один или более тирозинкиназных рецепторов.
Генетический контроль детерминации демонстрируется также существованием так называемых гомейотропных или гомеозисных мутаций, которые, как показано у насекомых, вызывают изменения при детерминации в специфических имагинальных дисках. В результате некоторые части тела развиваются не на своих местах. Например, у дрозофил мутации трансформируют детерминацию антенного диска в диск, который развивается в аппендикс конечности, протянутой от головы. У насекомых из рода Ophthalmoptera структуры крыльев могут развиваться из диска для глаз. У мышей показано существование генного кластера (комплекса) Нох, который состоит из 38 генов и контролирует развитие конечностей.
Самостоятельное значение имеет вопрос о регуляции активности генов в период эмбрионального развития. Считают, что в ходе дифференцировки гены действуют в разное время, что выражается в транскрипции в разных дифференцированных клетках разных мРНК, т. е. имеет место репрессия и дерепрессия генов. Например, количество генов, транскрибируемых в РНК в бластоцитах морского ежа, равно 10%, в клетках печени крыс — тоже 10%, а в клетках тимуса крупного рогатого скота — 15%. Предполагают, что в контроле транскрипционного статуса генов принимают участие негистоновые белки. В пользу этого предположения свидетельствуют следующие данные. Когда хроматин клеток в фазе транскрибируется в системе in vitro, то синтезируется только гистоновая мРНК, а вслед за нею и гистоны. Напротив, когда используют хроматин клеток из О^-фазы, то никакой гистоновой мРНК не синтезируется. Когда же негистоновые белки удаляются из хроматина G1-фазы и замещаются негис-тоновыми хромосомными белками, синтезированными в фазе S, то после транскрипции такого хроматина in vitro синтезируется гистоновая мРНК. Больше того, когда негистоновые белки происходят из G^-фазы клеток, а ДНК и гистоны из S-фазы клеток, никакой гистоновой мРНК не синтезируется. Эти результаты показывают, что содержащиеся в хроматине негистоновые белки определяют возможность транскрипции генов, кодирующих гистоны. Поэтому считают, что негистоновые хромосомные белки могут играть важную роль в регуляции и экспрессии генов у эукариот.
Имеющиеся данные позволяют считать, что в регуляции транскрипции у животных принимают участие белковые и стероидные гормоны. Белковый (инсулин) и стероидные (эстрогон и тестосте-рон) гормоны представляют собой две сигнальные системы, используемые в межклеточных коммуникациях. У высших животных гормоны синтезируются в специализированных секреторных клетках. Освобождаясь в кровяное русло, они поступают в ткани, поскольку молекулы белковых гормонов имеют относительно крупные размеры, то они не проникают в клетки. Поэтому их эффекты обеспечиваются белками-рецепторами, локализованными в мембранах клеток-мишеней, и внутриклеточными уровнями циклического АМФ (цАМФ). Напротив, стероидные гормоны являются малыми молекулами, вследствие чего легко проникают в клетки через мембраны. Оказавшись внутри клеток, они связываются со специфическими рецепторными белками, которые имеются в цитоплазме только клеток-мишеней. Как считают, комплексы гормон + белковый рецептор, концентрируясь в ядрах клеток-мишеней, активирует транскрипцию специфических генов через взаимодействие с определенными негистоновыми белками, которые связываются с промоторными районами специфических генов. Следовательно, связывание комплекса гормон + белок (белковый рецептор) с негистоновыми белками освобождает промоторные районы для движения РНК-полимеразы. Обобщая данные о генетическом контроле эмбрионального периода в онтогенезе организмов, можно заключить, что его ход контролируется дифференциальным включением и выключением действия генов в разных клетках (тканях) путем их дерепрессии и репрессии.
Постэмбриональный период
После появления организма на свет начинается его постэмбриональное развитие (постнатальное для человека), которое у разных организмов протекает от нескольких дней до сотен лет в зависимости от их видовой принадлежности. Следовательно, продолжительность жизни — это видовой признак организмов, не зависящий от уровня их организации (см. ниже).
В постэмбриональном онтогенезе различают ювениальный и пу-бертатный периоды, а также период старости, заканчивающийся смертью.
Ювенильный период. Этот период (от лат. juvenilis — юный) определяется временем от рождения организма до полового созревания. У разных организмов он протекает по-разному и зависит от типа онтогенеза организмов. Для этого периода характерно либо прямое, либо непрямое развитие.
В случае организмов, для которых характерно прямое развитие (многие беспозвоночные, рыбы, пресмыкающиеся, птицы, млекопитающие, человек), вылупившиеся из яйцевых оболочек или новорожденные сходны со взрослыми формами, отличаясь от последних лишь меньшими размерами, а также недоразвитием отдельных органов и несовершенными пропорциями тела (рис. 94).

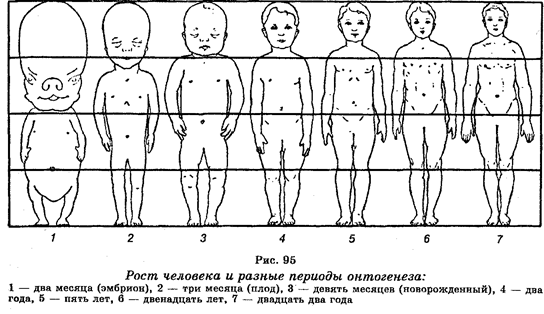
Характерной особенностью роста в ювенильный период организмов, подверженных прямому развитию, является то, что происходит увеличение количества и размеров клеток, изменяются пропорции тела. Рост человека в разные периоды его онтогенеза показан на рис. 95. Рост разных органов человека неравномерен. Например, рост головы заканчивается в детстве, ноги достигают пропорциональной величины примерно к 10 годам. Наружные половые органы очень быстро растут в возрасте 12—14 лет. Различают определенный и неопределенный рост. Определенный рост характерен для организмов, которые к определенному возрасту прекращают свой рост, например, насекомые, млекопитающие, человек. Неопределенный рост характерен для организмов, которые растут всю жизнь, например, моллюски, рыбы, земноводные, рептилии, многие виды растений.
В случае непрямого развития организмы претерпевают превращения, называемые метаморфозами (от лат. metamorphosis — превращение). Они представляют собой видоизменения организмов в процессе развития. Метаморфозы широко встречаются у кишечно-полостных (гидры, медузы, коралловые полипы), плоских червей (фасциолы), круглых червей (аскариды), моллюсков (устрицы, мидии, осьминоги), членистоногих (раки, речные крабы, омары, креветки, скорпионы, пауки, клещи, насекомые) и даже у некоторых хордовых (оболочечники и земноводные). При этом различают полные и неполные метаморфозы. Наиболее выразительные формы метаморфозов наблюдают у насекомых, которые подвергаются как неполным, так и полным метаморфозам.
 Неполное превращение — это такое развитие, при котором из яйцевых оболочек выходит организм, строение которого сходно со строением взрослого организма, но размеры намного меньше. Такой организм называют личинкой. В процессе роста и развития размеры личинок увеличиваются, но имеющийся хитипизированный покров мешает дальнейшему увеличению размеров тела, что приводит к линьке, т. е. сбрасыванию хитинизированного покрова, под которым находится мягкая кутикула. Последняя расправляется, и это сопровождается увеличением размеров животного. После нескольких линек животное достигает зрелости. Неполное превращение характерно, например, в случае развития клопов (рис. 96).
Неполное превращение — это такое развитие, при котором из яйцевых оболочек выходит организм, строение которого сходно со строением взрослого организма, но размеры намного меньше. Такой организм называют личинкой. В процессе роста и развития размеры личинок увеличиваются, но имеющийся хитипизированный покров мешает дальнейшему увеличению размеров тела, что приводит к линьке, т. е. сбрасыванию хитинизированного покрова, под которым находится мягкая кутикула. Последняя расправляется, и это сопровождается увеличением размеров животного. После нескольких линек животное достигает зрелости. Неполное превращение характерно, например, в случае развития клопов (рис. 96).
Полное превращение — это такое развитие, при котором из яйцевых оболочек освобождается личинка, существенно отличающаяся по строению от взрослых особей. Например, у бабочек и многих насекомых личинками являются гусеницы. Гусеницы подвержены линьке, причем могут линять по нескольку раз, превращаясь затем в куколки. Из последних развиваются взрослые формы (имаго), которые не отличаются от исходных (рис. 97).
У позвоночных метаморфозы встречаются среди земноводных и костных рыб. Для личиночной стадии характерно наличие провизорных органов, которые либо повторяют признаки предков, либо имеют явно приспособительное значение. Например, для головастика, являющегося личиночной формой лягушки и повторяющего признаки исходной формы, характерны рыбообразная форма, наличие жаберного дыхания, одного круга кровообращения. Приспособительными признаками головастиков являются их присоски, длинный кишечник. Для личиночных форм характерно также и то, что по сравнению со взрослыми формами, они оказываются приспособленными к жизни в совершенно иных условиях, занимая другую экологическую нишу и другое место в цепи питания. Например, личинки лягушек имеют жаберное дыхание, тогда как взрослые формы — легочное. В отличие от взрослых форм, которые являются плотоядными существами, личинки лягушек питаются растительной пищей.
Последовательность событий в развитии организмов часто называют жизненными циклами, которые могут быть простыми и сложными. Наиболее простые циклы развития характерны, например, для млекопитающих, когда из оплодотворенной яйцеклетки развивается организм, который снова продуцирует яйцеклетки и т. д. Сложными биологическими циклами являются циклы у животных, для которых характерно развитие с метаморфозами. Знания о биологических циклах имеют практическое значение, особенно в случаях, когда организмы являются возбудителями или переносчиками возбудителей болезней животных и растений.
Развитие и дифференциация, связанные с метаморфозами, являются результатом естественного отбора, благодаря которому многие личиночные формы, например, гусеницы насекомых и головастики лягушек адаптированы к среде лучше, чем взрослые половозрелые формы.
Пубертатный период. Этот период называют еще зрелым, и он связан с половой зрелостью организмов. Развитие организмов в этот период достигает максимума.
На рост и развитие в постэмбриональный период большое влияние оказывают факторы среды. Для растений решающими факторами являются свет, влажность, температура, количество и качество питательных веществ в почве. Для животных первостепенное значение имеет полноценное кормление (наличие в корме белков, углеводов, липидов, минеральных солей, витаминов, микроэлементов). Важны также кислород, температура, свет (синтез витамина Д).
Рост и индивидуальное развитие животных организмов подвержены нейрогуморальной регуляции со стороны гуморальных и нервных механизмов регуляции. У растений обнаружены гормонопо-добные активные вещества, получившие название фитогормонов. Последние влияют на жизненно важные отправления растительных организмов.
 В клетках животных в процессе жизнедеятельности синтезируются химически активные вещества, влияющие на процессы жизнедеятельности. Нервные клетки беспозвоночных и позвоночных вырабатывают вещества, получившие название нейросекретов. Железы эндокринной, или внутренней, секреции также выделяют вещества, которые получили название гормонов. Эндокринные железы, в частности, те, которые имеют отношение к росту и развитию, регулируются нейросекретами. У членистоногих регуляция роста и развития очень хорошо показана на примере влияния гормонов на линьку. Синтез личиночного секрета клетками регулируется гормонами, накапливающимися в мозге. В особой железе у ракообразных вырабатывается гормон, тормозящий линьку. Уровни этих гормонов определяют периодичность линек. У насекомых установлена гормональная регуляция созревания яиц, протекание диапаузы.
В клетках животных в процессе жизнедеятельности синтезируются химически активные вещества, влияющие на процессы жизнедеятельности. Нервные клетки беспозвоночных и позвоночных вырабатывают вещества, получившие название нейросекретов. Железы эндокринной, или внутренней, секреции также выделяют вещества, которые получили название гормонов. Эндокринные железы, в частности, те, которые имеют отношение к росту и развитию, регулируются нейросекретами. У членистоногих регуляция роста и развития очень хорошо показана на примере влияния гормонов на линьку. Синтез личиночного секрета клетками регулируется гормонами, накапливающимися в мозге. В особой железе у ракообразных вырабатывается гормон, тормозящий линьку. Уровни этих гормонов определяют периодичность линек. У насекомых установлена гормональная регуляция созревания яиц, протекание диапаузы.
У позвоночных железами внутренней секреции являются гипофиз, эпифиз, щитовидная, паращитовидная, поджелудочная, надпочечники и половые железы, которые тесно связаны одна с другой (рис. 98). Гипофиз у позвоночных вырабатывает гонадотропный гормон, стимулирующий деятельность половых желез. У человека гормон гипофиза влияет на рост. При недостатке развивается карликовость, при избытке — гигантизм. Эпифиз продуцирует гормон, который влияет на
сезонные колебания в половой активности животных. Гормон щитовидной железы влияет на метаморфоз насекомых и земноводных. У млекопитающих недоразвитие щитовидной железы ведет к задержке роста, недоразвитию половых органов. У человека из-за дефекта щитовидной железы задерживается окостенение, рост (карликовость), не наступает полового созревания, останавливается психическое развитие (кретинизм). Надпочечники продуцируют гормоны, оказывающие влияние на метаболизм, рост и дифференци-ровку клеток. Половые железы продуцируют половые гормоны, которые определяют вторичные половые признаки. Удаление половых желез ведет к необратимым изменениям ряда признаков. Например, у кастрированных петухов прекращается рост гребня, теряется половой инстинкт. Кастрированный мужчина преобрета-ет внешнее сходство с женщиной (не растет борода и волосы на коже, отлагается жир на груди и в области таза, сохраняется тембр голоса и т. д.).
Фитогормонами растений являются ауксины, цитокинины и гиб-береллины. Они регулируют рост и деление клеток, образование новых корней, развитие цветков и другие свойства у растений.
На всех периодах онтогенеза организмы способны к восстановлению утраченных или поврежденных частей тела. Это свойство организмов носит название регенерации, которая бывает физиологической и репаративной.
Физиологическая регенерация — это замена утерянных частей тела в процессе жизнедеятельности организма. Регенерации этого типа очень распространены в животном мире. Например, у членистоногих она представлена линькой, которая связана с ростом. У рептилий регенерация выражается в замещении хвоста и чешуи, у птиц — перьев, когтей и шпор. У млекопитающих примером физиологической регенерации может быть ежегодное сбрасывание оленями рогов.
Репаративная регенерация — это восстановление части тела организма, отторгнутой насильственным путем. Регенерация этого типа возможна у многих животных, но ее проявления различны. Например, она часта у гидр и связана с размножением последних, поскольку из части регенерирует весь организм. У других организмов регенерации проявляются в виде способности отдельных органов к восстановлению после утраты ими какой-либо части. У человека достаточно высокой регенеративной способностью обладают эпителиальная, соединительная, мышечная и костная ткани.
Растения многих видов также способны к регенерации. Данные о регенерации имеют большое значение не только в биологии. Их широко используют в сельском хозяйстве, в медицине, в частности, в хирургии.
Старость как этап онтогенеза. Старость является предпоследним этапом онтогенеза животных, причем ее длительность определяется общей продолжительностью жизни, которая является видовым признаком и которая у разных животных является разной. Наиболее точно старость изучена у человека.
Известны самые различные определения старости человека. В частности, одно из наиболее популярных определений заключается в том, что старость есть накопление последовательных изменений, сопровождающих повышение возраста организма и увеличивающих вероятность его болезней или смерти. Науку о старости человека называют геронтологией (от греч. geron — старик, старец, logos — наука). Ее задачей является изучение закономерностей возрастного перехода между зрелостью и смертью.
Научные исследования в геронтологии распространяются на разные области, начиная с исследований изменения активности клеточных ферментов и заканчива
|
|
|
