 |


Анатомические особенности строения вечнозеленых и листопадных психрофитных кустарничков семейства вересковые
|
|
|
|
Общие анатомические признаки у вересковых проявляются, прежде всего, в строении стебля. Так нами изучались строения стебля голубики, багульника и болотного мирта. Все они отличаются общими особенностями в строение стебля, которые заключаются, прежде всего, в следующем:
● стебель имеет четко выраженные три блока – первичная кора, центральный цилиндр и сердцевина;
● первичная кора образована толстостенными клетками колленхимы, где располагаются хорошо выраженные крупные воздухоносные полости;
● флоэма и ксилема представлены сплошными кольцами, т.е. стебель имеет типичное непучковое строение с кольцевым способом заложения камбия;
● сердцевина достаточно крупная, образована паренхимными клетками.
Снаружи стебель багульника покрыт однослойной покровной тканью – эпидермой – клетки ее имеют несколько вытянутую форму, плотно сомкнуты между собой. Далее располагается механическая ткань (предположительно уголковая колленхима) с сильно утолщенными клеточными стенками и малыми внутренними полостями. Между клетками первичной коры имеются достаточно крупные воздухоносные полости. Центральный цилиндр представлен многослойной флоэмой и трехслойной, в нашем случае исследовался трехлетний побег, ксилемой. Ксилема образована толстостенными прозенхимными клетками с достаточно сильно утолщенными клеточными стенками. В поперечном сечении форма клеток близка к прямоугольнику. Между клетками ксилемы и флоэмы располагается камбий, образующий вторичные проводящие ткани. Центральное положение занимает сердцевина, имеющая округлую форму и состоящая из очень крупных паренхимных клеток с неутолщенными клеточными стенками. Ее клетки выполняют запасающую функцию (рис.1.).
|
|
|
Стебель голубики имеет свои анатомические особенности, отличающие его от выше описанного стебля багульника (рис.2).
Эпидерма так же однослойная, но клетки более мелкие, чем у стебля багульника. Далее закладывается механическая ткань, уголковая колленхима с утолщенными клеточными оболочками и более крупными внутренними полостями, по сравнению с клетками стебля багульника. Воздухоносные полости примыкают непосредственно к флоэме, в отличие от стебля багульника, где они были полностью окружены механической тканью.
Флоэма состоит не только из ситовидных трубок, но включает в себя и лубяные волокна. В ксилеме хорошо выражены сердцевинные лучи, идущие от флоэмы к сердцевине стебля. Сердцевина имеет форму пятиугольника и образована клетками правильной паренхимной формы, имеющими небольшие межклетники.
Стебель болотного мирта снаружи покрыт однослойной эпидермой, клетки колленхимы утолщены незначительно, соответственно увеличиваются их внутренние полости. Воздухоносные полости в первичной коре самые крупные из всех изученных нами стеблей, близки по форме к прямоугольникам. Флоэма слабо развита, ситовидные трубки почти не просматриваются, в основном, она состоит из клеток лубяных волокон. Зато слой ксилемы развит достаточно мощно, хорошо выражены сердцевинные лучи, паренхима сердцевины одревесневшая, образована крупными плотносомкнутыми клетками (рис.3).

Рис.1. Анатомическое строение стебля багульника болотного (Ledum palustre): 1 – эпидерма; 2 – уголковая колленхима; 3 – воздухоносные полости; 4 – флоэма; 5 – вторичная ксилема; 6 – первичная ксилема; 7 – сердцевина; 8 – камбий.
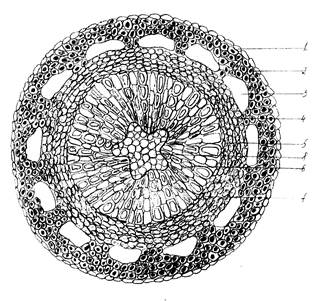
Рис.2. Анатомическое строение стебля голубики обыкновенной (Vaccinium uliginosum L.): 1 – эпидерма; 2 – колленхима; 3 – воздухоносные полости; 4 – флоэма; 5 – камбий, 6 – сосуды ксилемы; 7 – сердцевинные пучки; 8 – сердцевина.
|
|
|
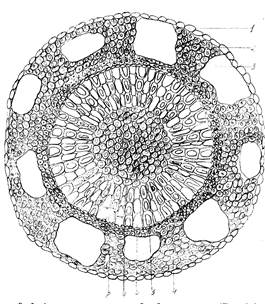
Рис. 3. Анатомическое строение стебля болотного мирта (Chamaedaphne calyculata): 1 – эпидерма; 2 – колленхима; 3 – воздухоносные полости; 4 –флоэма; 5 – ксилема; 6 – сердцевинный луч; 7 – сердцевина; 8 – камбий.
В общих чертах анатомическое строение стеблей вересковых соответствует условиям их произрастания – переувлажненным местам обитания с холодными почвами, что способствует развитию воздухоносных полстей. Поэтому они имеют признаки ксероморфной структуры. Это проявляется и в сплошном заложении проводящей ткани и в развитии механической ткани.
В анатомическом строении листьев растений семейства вересковых так как и стебля имеется значительное разнообразие, так встречаются вечнозеленые и листопадные виды вересковых.
Среди разнообразия листьев вересковых выделяют 3 типа:
● тип рододендрона – лист плоский или со слегка завернутыми вниз краями.
Такой лист имеют клюква, брусника, голубика и др.;
● игловидный лист – напоминает хвоинку как у некоторых видов эрики (Erica),
хариманеллы (Harrimanella);
● эрикоидный лист с сильно завернутыми книзу краями (у вереска).
У некоторых вересковых листья вообще уникальны. Так, кассиопея Редовского (C.redowskii) имеет лист, напоминающий палец перчатки, т.е. с почти замкнутой полостью. У многих вересковых лист может свертываться при наступлении засушливого периода; это отмечено, в частности, для брусники (Тимошок Е.Е., Паршина Н.В., 1992).
Таким образом, у ряда вересковых существуют динамические реакции на изменения погодных условий. Чрезмерное испарение воды в сухую погоду ослабляет напряжение в слоях мезофилла на нижней стороне листа, поэтому лист выгибается, скручивается; контакт листовой полости с окружающей средой уменьшается. Имеется, однако, сведения, что у очень многих вересковых, в частности у самого вереска, устьица есть и на верхней стороне листа. Таким образом, испарение происходит даже при полностью закрытой листовой полости. Значение этой полости становится понятным, если лист опустить в воду.
Анатомическое изучение листьев этих растений обнаруживает, что и "ксероморфность" их далеко не так ярко выражена как кажется.
Нами были изучены строение листьев некоторых психрофитных вечнозеленых и листопадных кустарничков. Так, например, при рассмотрении поперечного среза листа багульника мы обнаружили, что эпидерма однослойная, образована мелкими, плотно сомкнутыми клетками, на внешней стороне, контактирующей с атмосферой, откладывается толстый слой кутикулы. На нижней поверхности листа расположено большое количество простых и головчатых железистых волосков, устьица здесь не углубленные, а поверхностные. Число устьиц на единицу поверхности невелико.
|
|
|
Мезофилл листа гомогенный, имеет сильно развитую губчатую ткань, среди клеток которой находится большое количество межклетников. Сеть жилок не густая, проводящие ткани пучков развиты не сильно. Механических тканей, столь характерных дл листьев ксерофитов, почти нет (рис.4).
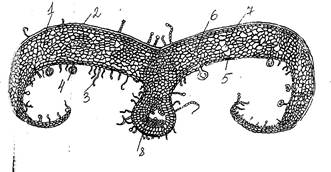
Рис.4 Поперечный срез листа багульника болотного (Ledum palustre): 1 – эпидерма; 2 – кутикула; 3 – кроющие волоски; 4 – железистые волоски; 5 – устьице; 6 – губчатый мезофилл; 7 – межклетники; 8 – закрытый коллатеральный проводящий пучок.
Также нами изучено строение листьев клюквы, относящейся к группе вечнозеленых кустарничков и голубики, принадлежащей к листопадным кустарничкам. В результате анатомических исследований мы выяснили, что анатомическое строение данных листьев имеет достаточное количество общих черт:
● Эпидерма однослойная, на внешней стороне верхней эпидермы имеет толстый защитный слой кутикулы;
● Устьица располагаются только на нижней стороне листа, среди клеток нижней эпидермы и на одном уровне с ее клетками, т.е. не являются погруженными;
● Мезофилл гетерогенный, четко дифференцирован на хорошо развитую палисадную ткань, как правило, двухслойную и губчатую;
● В губчатом мезофилле достаточное количество крупных межклетников и воздухоносных полостей, примыкающих к устьичным аппаратам.
Эпидерма четко подразделяется на нижний слой, состоящий из более мелких клеток, среди которых находятся устьица и верхний образованный крупными плотносомкнутыми клетками. Отличия же заключаются в том, что клетки эпидермы у листа голубики мельче, чем у листа клюквы, у которой отмечен и достаточно мощный слой кутикулы. Клетки мезофилла наоборот крупнее у листа голубики, палисадный мезофилл состоит из вытянутых клеток расположенных в два слоя, клетки губчатой ткани также имеют вытянутую форму, образуют межклетники и располагаются перпендикулярно нижней эпидерме (рис.5).
|
|
|

Рис.5. Поперечный срез листа голубики обыкновенной (V. Uliginosum L.): 1 – верхняя эпидерма; 2 – нижняя эпидерма; 3 – устьица; 4 – палисадный мезофилл; 5 – губчатый мезофилл; 6 – межклетники; 7 – воздухоносные полости.
Далее в своей работе мы изготовили поперечные срезы с листьев однолетнего и прошлогоднего листа клюквы и сравнили их строение (рис.6).
При изучении полученных микропрепаратов мы обнаружили, что в однолетнем листе клетки эпидермы мельче, а слой кутикулы тоньше. У листа второго года жизни эпидермальные клетки, особенно входящие в состав нижней эпидермы, значительно крупнее, а слой кутикулы толще почти в два раза. Мезофилл в первом случае слабо дифференцирован на слои палисадной и губчатой ткани и, в основном, представлен рыхлой губчатой тканью с достаточно крупными межклетниками. Лист второго года жизни имеет гетерогенный мезофилл, четко подразделяющийся на верхний палисадный слой, образованный двумя рядами клеток и нижний губчатый с межклетниками и воздухоносными полостями.
Все особенности строения изученных нами листьев не позволят считать их листьями типичного ксерофита: у них есть признаки мезоморфной и даже гигроморфной структуры. Таким образом, эта группа растений по своему анатомическому строению ближе к мезофитам.
Достаточное водоснабжение, но слабое минеральное питание, плохо аэрируемая и холодная почва вызывают общую задержку их роста, а произрастание на открытых, не защищенных от ветра зимой и хорошо освещаемых летом местах, привело к образованию у них мощной кутикулы, играющей роль экрана, отражающего яркие солнечные лучи и уменьшающего испарение. Интенсивным освещением можно объяснить и сворачивание листьев, которое уменьшает транспирацию.
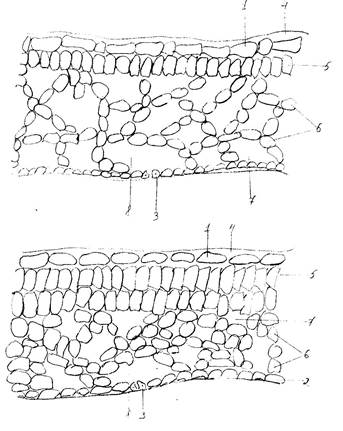
Рис.6. Поперечный срез листа клюквы обыкновенной (Oxycoccus
macrocarpus): 1 – верхняя эпидерма; 2 – нижняя эпидерма; 3 – устьица; 4 – кутикула; 6 – губчатый мезофилл; 7 – межклетник; 8 – воздухоносные полости.
ЗАКЛЮЧЕНИЕ
Специфика природных условий Севера и, в частности, экстремальные значения целого ряда жизненно важных для растений экологических факторов не могли не сказаться (и заметно сказались!) на морфологических, эколого-физиологических и биологических признаках, свойствах и особенностей северных растений. По совокупности данных показателей выделили отдельную экологическую группу растений – это растения психрофиты.
|
|
|
К ним относят виды, произрастающие в тайге, тундре, на высокогорьях. Сюда же могут относиться растения, живущие в условиях холодного, плохо прогревающегося даже летом, сфагнового болота.
Форма растений психрофитов многообразны и зависят как от природы организмов, так и от разностороннего воздействия окружающей среды.
Среди психрофитных растений встречаются представители семейств: осоковые, гвоздичные, крестоцветные, сложноцветные, большое количество видов семейства вересковые. Большинство вересковых – это вечнозеленые и листопадные кустарнички тундры и сфагновых болот.
Растения тундры живут в очень суровых условиях, что накладывает особый отпечаток на их облик. Большей частью – это приземистые или стелющиеся кустарнички с мелкими, обычно вечнозелеными, кожистыми листьями. Кроме вечнозелености, общим для них является особое строение листьев: они мелкие и завернуты краями вниз так, что прикрывается нижняя поверхность листа с устьицами.
Листья, в основном, мелкие, эпидерма однослойная с блестящей толстой кутикулой, часто с густыми волосками на нижней стороне листа. Число устьиц на единицу поверхности невелико. Мезофилл листа, в ряде изученных нами случаев, – гетерогенный, иногда имеет только губчатую ткань с большим количеством межклетников. Сеть жилок негустая, проводящие ткани пучков развиты несильно, механических тканей почти нет.
Общее анатомические признаки у психрофитных растений семейства вересковых проявляются и в строении стебля. Все изученные нами стебли отличались общими особенностями:
● стебель имеет четко выраженные 3 блока: первичная кора, центральный цилиндр и сердцевина;
● первичная кора образована толстостенными клетками колленхимы, где располагаются хорошо выраженные крупные воздухоносные полости;
● флоэма и ксилема представлены сплошными кольцами, т.е. стебель имеет типичное непучковое строение, с кольцевым способом заложения камбия.
Библиография
1. Алексеев С.В., Беккер А.М. Изучаем экологию экспериментально. – СПб.: Феникс, 1993. – 201с.
2. Алексеев Ю.Е., Баландин С.А. Энциклопедия растений России. Растения тундры. – М.: Классикс Стиль, 2003. – 208с.
3. Атлас ареалов и ресурсов лекарственных растений СССР – М.: ГУГК, 1976. – 340с.
4. Бавтуто Г.А., Ерей Л.М. Практикум по анатомии и морфологии растений. – Минск: Новое знание, 2002. – 458 с.
5. Богданова Г.А., МуратовЮ.М., Брусника в лесах Сибири. – Новосибирск: Наука, 1978. – 178с.
6. Большой атлас природы России / Под ред. М.Сергеевой, С.Бровкина. – М.: Эгмонт Россия Лтд, 2003. – 644 с.
7. Боч М.С., Смагин В.А. Флора и ратительность болот Северо-запада России и Принципы их охраны. – СПб.: Гидрометеоиздат, 1993. – 224с.
8. Брусника: Морфология и анатомия. Фитоценотическая приуроченность. Урожайность. Хранение и переработка. Химический состав ягод / Юдина В.Ф., Белоногова Т.В., Колупаева С.Н. и др. – М.: Наука, 1986. – 73с.
9. Бумар Г.И., Таргонский П.Н. Возрастной состав ценопопуляций Vaccinium vitis-idaea L. в Полесском государственном заповеднике (УССР) / Ратительные ресурсы. – СПб.: ГУГК 1989. – Т.25, вып.3. – с.349-353.
10. Викторов В.П., Гуленькова М.А., Дорохина Л.Н. и др. Практикум по анатомии и морфологии растений. – М.: Академия, 2001.
11. В краю селькупов: Красноселькупский район: природа.Люди.Экономика. экология / кол.авторов. – Екатеринбург: Сред.-Урал.кн.изд-во, 2000. -200с.
12. Ипатов В.С., Кирикова Л.А. Фитоценология. – СПб.: С.-Петербургского ун-та, 1997. - 316с.
13. Колесникова Н.А. Школьный учебно-опытный участок-лаборатория исследовательской работы учащихся. – Тобольск: изд. ТГПИ, 2003. – 61 с.
14. Крылов А.Г. Жизненные формы лесных фитоценозов. – СПб.: Наука, 1984. – 189с.
15. Лакин Г.Ф. Биометрия: уч.пособие. – 4-е изд., перераб. И доп. – М.: Высшая школа.1990. – 351с.
16. Лотова Л.И. Морфология и анатомия высших растений: Учебное пособие. – Киев: Эдиториал, 2000.
17. Мазная Е.А. Влияние промышленных выбросов на состояние и структуру ценопопуляций Vaccinium myrtillus L. И Vaccinium vitis-idaea L. (Кольский п-ов) //. Растительные ресурсы. – 2001. – Т.37вып.3.
18. Медведев С.С. Физиология растений. – СПб., 2004. – 336 с.
19. Минаева В.Г. Лекарственные растения Сибири. – Новосибирск: Наука, 1991. – 496 с.
20. Пояркова А.И. Ericaceae // Флора СССР. Т.18 – М. – Л., 1952. – С. 22-107.
21. Работнов Т.А. Луговедение. – М.: МГУ, 1984. – 319с.
22. Работнов Т.А. Фитоценология. – М.: МГУ, 1978. – 383с.
23. Рябицев В.К., Плотников В.В., Смирнов Н.Г., Ольшванг В.Н., Богданов В.Д., Мухин В.А., Троценко Г.В., Пешкова Н.В. Природа Тюменского Севера. – Свердловск: Средне-уральское книжное издательство, 1991. – 432 с.
24. Смирнова О.В. Структура травяного покрова широколиственных лесов. – М.:Наука, 1987. – 205с.
25. Сокол А.П. Экологические шкалы болотных растений // Взаимосвязи компонентов лесных и болотных экосистем средней тайги Приуралья. – СПб.: Наука, 1980. = С.230-240.
26. Толмачев А.И. К истории возникновения и развития темнохвойной тайги. – М.: АН СССР, 1954.
Приложение 1

Рис.1. Цветки и тычинки вересковых
Цветки: 1 – хамедафна болотная (Chamaedaphne calyculata); 2 – клюква (Oxycoccus) Тычинки: 3 – зеновия припудренная (Zenobia pulverulenta); 4 – хамедафна болотная (Chamaedaphne calyculata); 5 – лиония кустарниковая (Lyonia fruticosa); 6 – черника тычиночная (Vaccinium stamineum); 7 – черника обыкновенная (Vaccinium myrtillis); 8 – эллиотия кистевидная (Elliotia racemosa); 9 – гаультерия лежачая (Gaultheria procumbens); 10 – пиерис пышноцветный (Pieris floribunda); 11 – оксидендрум древовидный (Oxydendrum arboretum): а- тычиночные придатки; б- поры.

Рис.2. Пыльцевые зерна цветков растений семейства вересковых
|
|
|
