 |


Универсальность индивидуальных различий
|
|
|
|
Индивидуальные различия вне всякого сомнения ограничены человеком как видом. Напротив, изменчивость животных индивидов можно наблюдать на всех участках шкалы видов животных. Конечно, поверхностное знакомство с ними производит впечатление близкого сходства и даже тождественности членов какой-либо группы животных. Но более близкое знакомство все-
Распределение индивидуальных различий 6 5
гда приводит к заключению, что между ними существуют индивидуальные различия. Для большинства людей все кошки, как им кажется, ведут себя «как кошки», но это мнение может измениться: для человека, имеющего любимое животное, каждая кошка, несомненно, обладает индивидуальностью.
Как можно догадаться, самые ранние сообщения об индивидуальных различиях среди животных содержали сведения о крайних случаях отклонений, «гениях» животного царства. Одним из наиболее известных и достоверных случаев являлся немецкий шеппард по кличке Приятель (30, 31). В отличие от других собак, Приятель наблюдался психологами в тщательно контролируемых условиях. Это животное научилось правильно реагировать на самые разные вербальные команды, имело ассоциации приблизительно на 400 слов, обозначающих конкретные объекты и действия. Особенно интересен тот факт, что Приятель адекватно реагировал даже тогда, когда одна и та же команда выражалась разными словами или когда одни и те же слова включались в состав разных команд. Кроме этого, было установлено, что он был в состоянии отвечать на слова, как таковые, даже тогда, когда жесты и другие вторичные подсказки устранялись с помощью экрана.
Можно привести и другие примеры собак, которых с успехом обучили решать запутанные задачи и отвечать на большое количество сложных команд. Наиболее прославились собаки-поводыри «Видящего глаза» и «Собачьего корпуса», которых эффективно использовали для военных целей во время обеих мировых войн. Шимпанзе обучались множеству специальных функций, таких как катание на коньках, езда на велосипеде, еда при помощи ножа и вилки, открывание дверей. Следует упомянуть также достижения цирковых животных всех видов, особенно «музыкальных» морских львов.
|
|
|
Со становлением сравнительной психологии и развитием контролируемых лабораторных измерений поведения животных, свидетельства об индивидуальных различиях вышли на другой уровень по сравнению с анекдотичными наблюдениями и отдельными исследованиями. Каждое лабораторное исследование, в котором было задействовано более одной особи, приводило к открытию индивидуальных различий. Специалисты по психологии животных, как правило, не интересуются измерениями из-
3 Дифференциальная психология
66 Дифференциальная психология
менчивости, поэтому данные по этой проблеме носят фрагментарный характер и часто не представлены в количественной форме. Но, когда бы ни приводились такие данные, границы их проявления в случайно отобранных группах на удивление широки. На каждой фазе исследуемого поведения фиксировалась большая индивидуальная изменчивость, например, в том, что касается общей спонтанной активности животных, относительной силы их влечений, эмоциональности, скорости перемещения, быстроты овладения навыками решения простых задач и поведения в более сложных проблемных ситуациях.
Аналогичным образом, индивидуальные различия наблюдались у каждого вида животных, от простейших одноклеточных организмов до человекообразных обезьян. В одном исследовании видов протозоа был найден, например, широкий спектр индивидуальных различий в скорости установления обусловленной реакции на свет. Процедура предполагала предъявление повторяющихся пар тактильных и световых стимулов. Число попыток, или повторов, необходимых для различных особей, находилось в пределах от 79 до 284, со средним значением 138,5 (ср. 21, с. 308). Индивидуальные различия были также отмечены среди парамеций по результатам научения их избегать тубу: некоторые экземпляры, например, так и не смогли этому научиться (8). В другой серии экспериментов на парамециях особи различались по склонности к образованию групповых масс или к свободному плаванию, а также по склонности к принятию решения входить или не входить им в питательный раствор, содержащий примеси посторонних химических веществ (7).
|
|
|
Не так давно Хирш и Трион (15) разработали остроумные техники для получения надежных измерений индивидуальных различий многочисленных поведенческих реакций простейших организмов. Разработанная ими процедура была применена, в частности, при исследованиях роли наследственности в поведении фруктовой мухи дрозофилы. Такое исследование будет рассмотрено в главе 4. Здесь же имеет смысл отметить, что уже на начальной стадии этих экспериментов был обнаружен широкий спектр индивидуальных различий. В одном таком исследовании мухам предоставлялось до 15 попыток, чтобы залететь в тестовую тубу, и результаты распределились от 0 до 15. Иными словами, некоторые мухи делали это каждый раз, а некоторые — ни-
Распределение индивидуальных различий 67
когда. Остальные особи распределились по всем значениям между этими двумя крайними членами.
Но самым популярным экспериментальным животным является белая крыса. Крыса, бегущая по лабиринту, с целью «просветить» психологов, стала символом современной психологии. Из всех экспериментов, демонстрирующих индивидуальные различия в научении крыс, мы упомянем только один. В эксперименте, который в более полном виде будет рассмотрен в главе 4, Трион (29) дал 142 крысам 19 попыток преодолеть лабиринт. Во время каждой попытки всякий раз, когда крыса забегала в тупик, это фиксировалось для подсчета ее ошибок. Общее число таких ошибок, совершенных во время осуществления 19 попыток, у разных индивидов колебалось в пределах от 7 до 214.
|
|
|
Довольно большой набор данных по индивидуальным различиям в обучении животных был получен в лаборатории Колумбийского университета в результате проведения в 30-х годах XX века серий исследовательских проектов по сравнительной психологии. Малые выборки гвинейских свиней, крыс-альбиносов, обыкновенных короткошерстных кошек и двух видов обезьян прошли через тестирование «проблемным ящиком», в котором животным предлагалось совершать все более сложные последовательности действий.
Ящик, изображенный на рисунке 21, состоит из внешней и внутренней клеток, последняя содержит в себе приманку, которую животное получает по завершении каждой успешной попытки. Во внутренней клетке расположены три дощечки, на которые животное должно в определенном порядке наступить, чтобы дверь в отделение с приманкой открылась. Задача первого «шага» состоит в том, чтобы наступить на первую дощечку, находящуюся справа от животного, зашедшего в коробку. Затем надо было наступить на дощечки 1 и 2; 1, 2 и 3; 1, 2, 3 и снова 2; 1,2, 3, 2, 1 и так далее в других комбинациях.
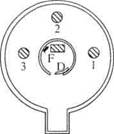
Е — входное отделение; D — дверь в отделение с едой; F — приманка; 1, 2, 3 — дощечки, на которые животное
Должно наступить
Рис. 21. Изображение проблемного ящика, использованного в исследованиях научения животных в Колумбийском университете. (Данные из Фьелда, 6, с. 403.)
68 Дифференциальная психология
Хотя исследования, проводимые с проблемным ящиком были направлены главным образом на то, чтобы выяснить, сколькими «шагами» могут овладеть животные каждого вида, полученные данные однозначно свидетельствовали о внутривидовых индивидуальных различиях. Индивиды различались между собой не только количеством попыток, которые были необходимы им для обучения каждому «шагу», но и общим количеством «шагов», которым они могли научиться. В таблице 2 приводятся сведенные вместе данные, включая крайние значения количества «шагов», которыми овладел каждый вид, число индивидов, обучившихся делать 1 «шаг», а также среднее значение, крайние значения и стандартные отклонения множества качеств, необходимых для обучения 1 «шагу». Последние данные касаются только 1 «шага», поскольку несколько членов в каждой группе смогли овладеть только им одним.
|
|
|
Таблица 2 Индивидуальные и групповые различия в обучении животных, измеренные при помощи проблемного ящика. (Данные из Фьелда, 6, с. 528 и Коха, 16, ее. 186, 208.)
| Кол-во | Кол-во | Попытки | обучиться | шагу 1 | ||
| Тип | Пределы | |||||
| животного | случаев | овладевших 1 шагом | В среднем | Пределы | СО | усвоенных шагов |
| Гвинейские | 185,50 | 53-407 | 176,28 | 0-1 | ||
| свиньи | ||||||
| Крысы- | 221,04 | 30-453 | 125,26 | 0-2 | ||
| альбиносы | ||||||
| Кошки | •46,69* | 9-136 | 25,28 | 3-7 | ||
| Обезьяны | 162,47 | 19-310 | 94,36 | 2-22 | ||
| (резус) | ||||||
| Обезьяны | 137,17 | 42-327 | 108,41 | 5-15 | ||
| (гебус) | ||||||
*В данном исследовании проблема, заключенная в шаге 1, для кошек была упрощена: животному разрешалось наступать на любую из трех дощечек. Поэтому данные по первому «шагу» в этой группе нельзя напрямую сравнивать с данными по другим видам.
Таблица 2 показывает, что среди гвинейских свиней некоторым так и не удалось обучиться даже первому «шагу», в то время как другие смогли это сделать; среди крыс некоторые обучились двум «шагам», некоторые одному, а некоторые ни одно-
Распределение индивидуальных различии 6 9
му; среди кошек значения лежат в пределах от 3 до 7; среди резус-обезьян — от 2 до 22 и среди гебус-обезьян — от 5 до 15. Таким образом, изменчивость оказалась столь велика, что среди представителей «высших» видов можно было легко найти тех, кто был не в состоянии обучиться тому, что удавалось некоторым представителям «низших» видов. Результаты покажутся нам еще более впечатляющими, если мы обратим внимание на число» попыток, необходимых для обучения первому «шагу». Например, гвинейская свинья и крыса, обладающие лучшими показателями, обучились этому шагу за меньшее число попыток, чем обезьяны обоих видов, имеющие худшие показатели. Такое частичное совпадение распределений, принципиально отличных друг от друга видов животных, является очень важным для всех групповых сравнений; мы будем часто возвращаться к нему, рассматривая исследования, проведенные с людьми.
Несмотря на то что большинство исследований животных было связано с той или иной формой научения, существуют аналогичные данные по индивидуальным различиям и в отношении других психологических характеристик. Что касается эмоциональных и мотивационных качеств, то исследования, проведенные на белых крысах, показали существование четких индивидуальных различий в проявлениях страха и агрессивности, в исследовательском поведении и в силе сексуального влечения (ср. 11). Исследования показали, что в проявлениях всех этих качеств существует «ситуационная общность», достаточная для того, чтобы позволить охарактеризовать поведение каждого индивида через призму данного качества. Например, крысы, тестировавшиеся в условиях четырех разных ситуаций, проявили тенденцию к постоянной агрессивности. Подобные результаты были получены и в отношении других качеств, упомянутых выше.
|
|
|
Получение иллюстративных данных по «высшим» видам предварялось изучением «темперамента» шимпанзе, описанного Геббом (13). Наблюдения проводились в ситуациях, позволявших шимпанзе по-разному реагировать на людей или на неживые предметы. Когда в классификации поведения животных для каждого типа поведения были использованы свои, тщательно подобранные категории — например, дружелюбность, агрессивность, избегание и т. д., — то обнаружился широкий спектр индивидуальных различий, которые можно было измерить с дос-
70 Дифференциальная психология
таточной надежностью. Ниссен (19) предоставил данные о больших индивидуальных различиях среди 57 шимпанзе, живущих в Йоркской лаборатории биологии приматов, расположенной в Оранж-парке (Флорида). Заметные расхождения наблюдались почти во всех поведенческих функциях, включая быстроту, склонность к чистоте, предпочтения в еде и связанные с этим привычки, эмоциональную экспрессивность, возбуждаемость, скорость и точность при обучении различным задачам, в использовании предметов и социальном взаимодействии с другими шимпанзе и людьми.
Приведенные иллюстрации достаточны для того, чтобы считать индивидуальные различия неотъемлемой, доступной для измерения, принадлежностью всего живого. Точные конфигурации, показывающие изменчивость, значения не имеют. Они не в состоянии дать нам абсолютное определение степени изменчивости у различных видов. Они также не позволяют нам производить общие сравнения, которые превосходили бы сравнение людей по разным качествам. Конкретные результаты выражаются в единицах измерения, специфических для каждого теста или других исследовательских процедур. Полученные данные, однако, подразумевают, что представители одного и того же вида различаются между собой своими поведенческими особенностями и что такие различия достаточно велики, поэтому распределение представителей одного вида частично совпадает с распределением индивидов совершенно других видов.
РЕЗЮМЕ
Индивидуальные различия носят количественный, а не качественный характер. Различия между индивидами выражаются соответствующей степенью. Частотные распределения для основных качеств показывают тяготение большинства индивидов к центру ряда и постепенное снижение их числа при приближении к крайним значениям. Такое распределение графически может быть представлено как в виде полигона частот, так и в виде гистограммы. Оно может быть в дальнейшем дополнено вычислениями центральной тенденции (среднего арифметического, медианы или моды) и измерениями изменчивости, такими как
Распределение индивидуальных различий 71
вычисление крайних значений стандартных отклонений. Многие графики распределения физических или психологических качеств приближаются к графику нормального распределения.
Распределения, которые отличаются от нормального графика скошенностью, остротой или другими характеристиками, могут получаться по многим причинам. Среди главных факторов, воздействующих на форму графика распределения, можно назвать следующие: неправильно составленная выборка; характерные особенности средств измерения, такие как неадекватность уровня сложности и несовершенство единиц измерения; особые условия, влияющие на сами измеряемые качества. Последние рассматривались в связи с гипотезой J-кривой социального конформизма, распределением редких событий Пуассона и возрастанием частоты на отрезке низких значений ряда вследствие действия патологических условий.
Абсолютизирующие утверждения в отношении степени изменчивости различных качеств являются бессмысленными по нескольким причинам, среди которых главные: несравниваемые единицы измерения и отсутствие абсолютной нулевой точки на психологических шкалах. Степень индивидуальных различий внутри специфических групп по конкретным свойствам может быть определена для практических целей в рамках соответствующего контекста. Но не имеет большого смысла делать общие утверждения о пределах человеческой изменчивости в отношении различных качеств.
Индивидуальные различия универсальны и для животных. Любое исследование поведения животных от простейших одноклеточных организмов до человекообразных обезьян открывает широкий спектр индивидуальных различий. Эти различия настолько велики, что наблюдается частичное совпадение распределений индивидов, относящихся к удаленным друг от друга видам.
БИБЛИОГРАФИЯ
1. Allport, F. H. The J-curve Hypothesis of conforming behavior. / soc. Psychol., 1934, 5, 141-183.
2. Anastasi, Anne. Practice and variability. Psychol. Monogr., 1934, 45, No. 5.
72 Дифференциальная психология
3. Blum, M. L. Industrial psychology and its social foundations. (Rev. Hd.) N.Y.: Harper, 1956.
4. Boring, E. G. A history of experimental psychology. (Rev. Ed.) N.Y.: Appleton-Cenhiry-Crofts, 1950.
5. Cook, W. W. Individual-trait differences in public schools with implications for school organization and curriculum development. Teach. Coll. J., 1947, 1956-57, 67-70.
6. Fjeld, Harriet A. The limit: of learning ability in rhesus monkeys. Genet. Psych. Monogr., 1934, 15, 369-537.
7. French, J. W. Individual differences in Paramecium. /. сотр. Psychol., 1940, 30.451-456.
8. French, J. W. Trial and error learning in Paramecium. /. exp. Psychol., 1940, 26, 609-613.
9. Chiselli, E. E., and Brown. C. W. Personnel and industrial psychology. N.Y.: McCraw-Hil, 1955.
10. Guillford, J. P. Fudamental statistics in psychology and education. (3rd Ed.) N.Y.: McGraw-Hill, 1956.
11. Hall, С S. Individual differences. In С. Р. Stone (Ed.), Comparative psychology. (3rd Ed.) N.Y: Prentice-Hall, 1951, Pp. 363-387.
12. Harris, J. A., Jackson, C. M., Paterson, D. G., and Scammon, R. L. The measurement of man. Minneapolis: Univer. Minn. Press, 1930.
13. Hebb, D. O. Temperament in chimpanzees: J. Method of analysis. / сотр.
physiol. Psychol., 1949, 42, 192-206.
14. Hildreth, Gertrude H. Individual differences. In W. S. Munroe (Ed.), Encyclopedia of educational research. (Rev. Ed.). NY.: Macmillan, 1950, 564-571.
15. Hirch, J., and Tryon, R. C. Mass screening and reliably individual measurement in the experimental behavior genetics of lower organisms. Psychol. Bull., 1956, 53, 402-410.
16. Koch, A. M. The limits of learning ability in cebus monkeys. Genet. Psychol. Monogr., 1935, 17. 165-234.
17. Laughlin, H. H. Racing capacity in the thoroughbred horse. Sci. Mon., 1934, 38, 210-222, 310-321.
18. Mintz, A., and Blum, M. A re-examination of the accident-proneness concept. /. одр/. Psychol, 1949, 33, 195-211.
19. Nissen, H. W. Individuality in the behavior of chimpanzees. Amer. Anthropologist. 1956, 58, 407-413.
20. Pepinsky, Pauline N. The J-eurve re-visited. /. abnorm. soc. Psychol, 1951, 46, 534-538.
21. Razran, G. Conditioned responses in animals other than dogs. Psychol.Bull, 1933, 30, 261-324.
22. Roberts, J. A. Fraser. The genetics of mental deficiency. Eugen. Rev., 1952,44, 71-83.
Распределение индивидуальных различий 7 3
23. Roberts, J. A. Fraser, Norman, R. M., and Criffiths, Ruth. Studies on a child population. IV. The form of the lower end of the frequency distribution of Stanford-Binet intelligence quotients and the fall of low intelligence quotients with advancing age. Ann. Eugen., 1938, 8, 319-330.
24. Ruggles, R., and Allport, G. W. Recent applications of the A-S Reaction Study. J. abnorm. soc. Psychol., 1939, 34, 518-528.
25. Scottish Council for Research in Education. The trend of Scottish intelligence. London: Univer. London Press, 1949.
26. Slovens, S. S. Mathematics, measurement, and psychophysies. In S. S. Stevens (Ed.). Handbook of experimental psychology. N.Y.: Wiley, 1951, 1-49.
27. Terman, L. M., and Merrill, Mand A. Measuring intelligence. Boston: Houghton Mifflin, 1937.
28. Tiffin, J., and Wirt, S. E. Determining visual standards for industrial jobs by statistical methods. Trans. Amer. Acad. Opthal. Otolaryng., 1945, 50, 72-93.
29. Tryon, R. C. Genetic differences in muze-learning ability in rats. 39fh Yearb., Nat. Soc. Stud. Educ, 1940, Part I, 111-119.
30. Warden, C. J. The ability of «Fellow,» lamous German shepherd dog. To understand language. J. genet. Psychol., 1928, 35, 330—331.
31. Warden, C. J., and Warner, L. H. The sensory capacities and intelligence
of dogs, with a report on the ability of the noted dog «Fellow» to respond to verbal stimuli. Quart. Rev. Biol., 1928, 3, 1—28.
32. Wechsler, D. The range of human capacities. (Rev. Ed.) Baltimore: Williams & Wilkins, 1952.
33. Wenger, M. A., and Ellington, M. The measurement of autonomic balance in children: method and normative data. Psychosom. Med., 1943, 5, 241-253.
34. Yule, G, U., and Kendall, M. G. An introduction to the theory of statistics. (13th Ed.) London: Griffin, 1919.
Глава 3
НАСЛЕДСТВЕННОСТЬ И СРЕДА:
ОСНОВНЫЕ ПОЛОЖЕНИЯ
Почему индивиды отличаются друг от друга? Какие факторы влияют на возникновение подобных отличий? Эти вопросы всегда вызывали длительные обсуждения и оживленные дискуссии. Кроме своей фундаментальной теоретической важности, проблема причинности индивидуальных различий имеет во многих областях далеко идущее практическое значение. Любая деятельность, направленная на улучшение поведенческих характеристик, должна основываться на понимании факторов, влияющих на изменения в поведении. Все методы в системе образования основываются на скрытых допущениях, имеющих прямое отношение к причинам индивидуальных различий. Точно так же ответы на многие вопросы, касающиеся свободы выбора и использования своих возможностей, групповых отношений и социальной структуры, зависят от знания причин психологических различий между индивидами разного возраста или пола, между этническими или национальными группами, между представителями разных социоэкономических уровней.
ПРИРОДА НАСЛЕДСТВЕННОСТИ1
Основу индивидуальных различий следует искать в наследственности индивида и в условиях окружающей среды, в которых он развивается. Давайте для начала рассмотрим то, что под-
1 Чтобы дополнить наш краткий экскурс в механизмы наследования, читатель должен обратиться к современным работам по генетике, таким, например, как книги Синнотта, Данна и Добжански (22), Снайдера и Дэвида
Наследственность и среда 75
разумевается под термином «наследственность». Естественно, что первой мыслью, возникающей при этом, является предположение, что понятие «наследственность» означает наследственность биологическую. Только фигурально мы можем говорить о «социальной наследственности», употребляя, например, выражение «культурное наследие двадцатого века» или «семейная удачливость». Так называемое социальное наследие в действительности не может устоять под влиянием окружающей среды.
Индивидуальная наследственность человека в основном состоит из особых генов, которые он получает от родителей при зачатии. Чтобы оказывать определенное влияние, фактор наследственности должен, таким образом, иметь особый ген или комбинацию генов. Гены группируются в хромосомы, или «окрашивающиеся тельца», названные так потому, что они становились видимы внутри клеточного ядра, когда окрашивались для наблюдения. Хромосомы соединены попарно, два члена каждой пары сходны между собой по своему проявлению и функции. Число хромосом в каждой клетке в целом постоянно для каждого вида, но отличается у разных видов. Каждая человеческая клетка, например, содержит 48 хромосом (24 пары); в каждой клетке комара их 6 (3 пары); а в каждой клетке рака их 200 (100 пар).
Хромосомы видимы под микроскопом, они похожи на прутики, колбаски или V-образные тельца (см. рис. 22). Однако гены внутри каждой хромосомы такие мелкие, что невидимы даже под сильным микроскопом. Через наблюдение за гигантскими хромосомами, которые были открыты внутри слюнных желез некоторых видов мух, стало возможным более полно при помощи микроскопа исследовать внутреннюю структуру хромосом. И хотя по оценкам они в 1000 или в 2000 раз больше
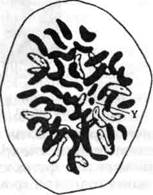
Рис. 22. Человеческие хромосомы под микроскопом; увеличение в 3600 раз. (Данные из Эванса и Суизи, 10.) (23) или Винчестера (26). Блестящий обзор современных достижений в генетике, подготовленный группой экспертов, можно найти у Данна (9). Более популярно обо всем этом можно прочесть у Шенфельда (21).

А. Фотография, сделанная под сильноувеличивающим микроскопом

В. Электронная микрография микроскопического сегмента с увеличением в 26000 раз
Рис. 23. Фрагмент гигантской хромосомы из слюнных желез фруктовой мухи. (Данные из Пейнтера, 18, с. 464 и Бейкера, 19, с. 9.)
других хромосом, по всем другим существенным характеристикам эти гигантские хромосомы подобны остальным хромосомам, найденным в других клетках тела. Рисунок 23 показывает сегмент такой гигантской хромосомы из слюнных желез фруктовой мухи Drosophila melanogaster (дрозофила меланогастер). Верхняя часть этого рисунка содержит фотографию, сделанную под сильноувеличивающим микроскопом; вторая часть содержит электронную микрографию с увеличением в 26000 раз. Частицы, видимые на микрографии, являются генами. Внутри одного и того же сегмента, или связки хромосомы, эти частицы выглядят одинаковыми по размеру и форме, но отличаются от частиц в других связках (19).
Каждый индивид начинает свою жизнь с одной клетки — это оплодотворенная яйцеклетка. Эта клетка делится на две дочерние клетки, каждая из которых снова делится и т. д. до тех пор, пока миллионы клеток не становятся зрелым организмом. Во время процесса клеточного деления, известного как митоз,
Наследственность и среда 77
каждая хромосома внутри клеточного ядра удваивается, разрываясь на две части, и каждая клетка, возникшая в результате такого деления, получает идентичный набор хромосом. Все клетки тела, таким образом, имеют одинаковую наследственность. Из одних клеток развиваются глазные клетки, из других — кожа, кости и т. д. в зависимости от влияния клеточного окружения. Такие условия, как сила тяготения, давление, пригодность кислорода и других химических веществ, а также электрические поля действуют по-разному на разные клетки, в зависимости от положения клетки по отношению к другим клеткам. С технической точки зрения это означает, что устанавливаются «дифференциальные градиенты» развития, такие как поверхностно-внутренние, дорсально-вентральные или антеро-постериорные градиенты. Следуя изначальной клеточной дифференциации, гены по-разному взаимодействуют с клеточным окружением, вызывая последующее специализированное развитие различных клеток.
Когда индивид достигает половой зрелости, другой тип клеточного деления приводит к формированию специализированных репродуктивных клеток, женских яйцеклеток или мужских сперматозоидов. Этот процесс называется мейозом, или уменьшением, поскольку число хромосом в каждой репродуктивной клетке в два раза меньше изначального. Вместо удвоения, происходящего при митозе, две хромосомы в каждой паре разделяются и расходятся по дочерним клеткам. Необходимо отметить, что при данном типе клеточного деления каждая клетка может получить разную комбинацию хромосом, поскольку хромосомы в каждой паре соединяются случайно. Более того, хромосомы не всегда разделяются, сохраняя целостность половинок в дочерних клетках, но сегменты одной половины могут соединяться с сегментами другой («кроссинговер»), увеличивая, таким образом, разнообразие возможных генных комбинаций в дочерних клетках. Когда яйцеклетка с набором материнских хромосом при зачатии оплодотворяется сперматозоидом с набором отцовских хромосом, общее число хромосом восстанавливается и сохраняется при последующем митозе развивающегося плода.
Наследственной основой индивидуальных различий является почти бесконечное разнообразие возможных генных комбинаций, особенно в таком сложном организме, как человек. Во-первых, необходимо отметить, что в целом даже простые
78 Дифференциальная психология
свойства человека зависят от совокупного влияния большого числа генов. Во-вторых, родительские клетки, принимающие участие в процессе зачатия, содержат различные комбинации генов, возникшие в результате мейоза. В-третьих, когда клетки двух организмов, материнского и отцовского, соединяются для создания нового организма, то тем самым они еще больше увеличивают разнообразие возможных генных комбинаций. Из этого следует, что двое детей от одних родителей, то есть братья или сестры, не могут иметь одинаковую наследственность. То же самое справедливо для разнояйцевых близнецов, которые хоть и рождаются в одно и то же время, но развиваются из разных яйцеклеток, и с точки зрения наследственности похожи друг на друга не более, чем обычные братья или сестры. Разнояйцевые близнецы могут быть одного пола, а могут быть и разнополыми, они могут быть не похожими друг на друга. С другой стороны, однояйцевые близнецы развиваются из разделившихся половинок единой оплодотворенной яйцеклетки, и поэтому имеют один и тот же набор генов. Такие индивиды с точки зрения наследственности являются копиями друг друга.
Простейшей иллюстрацией механизма наследственности могут служить единичные факторы, зависящие от одной-един-ственной пары генов. Примером такого единичного фактора является альбиносность, или отсутствие пигментации в глазах, волосах и на коже. Если индивид получил ген альбиносности от каждого из своих родителей (ее), он сам станет альбиносом. Индивиды с двумя генами нормальной пигментации (ее) будут иметь нормальную пигментацию. Оба этих индивида являются гомозиготными в отношении альбиносности. Это означает, что оплодотворенная яйцеклетка, или зигота, из которой такие индивиды выросли, получила одинаковые гены или альбиносности, или нормальной пигментации от обоих родителей. Если индивид получил ген альбиносности от одного родителя и ген нормальной пигментации от другого (Сс), он называется гетерозиготным по данному признаку. Такой индивид будет проявлять нормальную пигментацию, поскольку нормальная пигментация является доминирующим признаком, а альбиносность — рецессивным. Иными словами, альбиносность, будучи рецессивным признаком, проявляется только тогда, когда индивид получает рецессивный ген альбиносности от обоих родителей. Гетерозиготные индивиды
Наследственность и среда 79
(Сс), хотя и выглядят как люди с нормальной пигментацией, тем не менее несут в себе ген альбиносности, который они могут в свою очередь передать своим потомкам.
А другие единичные признаки не проявляют доминирования. Например, возьмем расцветку домашней птицы. Черная и пятнисто-белая птицы несут в себе генные пары, ни одна из которых не является доминантной. Их скрещивание даст потомков третьего цвета, так называемых «голубых андалузцев», не похожих ни на одного из родителей.
Необходимо добавить, что в случае, когда некие признаки по крайней мере в настоящее время рассматриваются как доминантно-рецессивные, дальнейшие исследования могут не обнаружить доминирования (ср. 6). Когда признак является доминантно-рецессивным, гетерозигота, естественно, не отличается от одной из гомозигот. Поэтому в действительности его гетерозиготность часто просто не берется в расчет. С развитием техники анализа, повышением степени ее доступности отличия могут быть найдены. Например, в случае патологии у носителя соответствующей гетерозиготы она может проявляться в слабой форме, которую можно обнаружить современными методами. Наличие гетерозиготности можно определить посредством химического анализа или других тонких технологий. Даже когда различие между гетерозиготностью и гомозиготностью не имеет практического значения в жизни индивида, это может быть полезной информацией, которую должны будут учитывать его потомки.
Что касается пола индивида, то он детерминируется парой хромосом, известных как половые хромосомы, которые обозначаются X и Y. Если ребенок получает Х-хромосому от каждого родителя, он будет женского пола; если получает хромосомы X и Y, то мужского. От своей матери ребенок может получить только X хромосомы, от отца — X, или Y. Y-хромосома сравнительно мала, считается, что в ней содержится очень мало генов. Такая Y хромосома изображена на рисунке 22. Современная генетика считает, что при определении пола действительно происходит взаимодействие генов Х-хромосомы с генами другой хромосомы. Существует мнение, что в каждом индивиде есть все необходимые для обоих полов гены. Но наличие двух Х-хромо-сом приводит к развитию женских половых признаков и подав-
80 Дифференциальная психология
лению мужских. Если же есть только одна Х-хромосома, то развитие получают мужские признаки.
Определенные гены в Х-хромосоме отвечают за признаки, связанные с полом. Среди наиболее известных — дальтонизм и гемофилия (интенсивное и продолжительное кровотечение из-за низкой сворачиваемости крови). Оба эти признака зависят от рецессивного гена, заключенного в Х-хромосоме. Если дочь наследует такой признак только от одного из родителей, то нормальный ген в другой Х-хромосоме, являющийся доминантным, не даст проявиться дефекту или заболеванию. Они проявятся только в том случае, если девочка унаследует несущий дефект ген от обоих родителей. Если же такой ген, содержащийся в Х-хромосоме, получает мальчик, дефект неизбежно проявится, поскольку в его Y-хромосоме нет гена, соответствующего нормальному развитию. Следовательно, подобные признаки более распространены среди мужчин, чем среди женщин.
Некоторые другие признаки, такие как облысение, являются зависимыми от пола, то есть они ведут себя как доминантные у одного пола, и как рецессивные — у другого. Таким образом, облысение будет развиваться у мужчины, если ген, связанный с облысением, был передан ему кем-либо из родителей. У женщины облысение будет развиваться, только если гены облысения были получены ею от обоих родителей. Еще одна группа признаков, известных как ограниченно-половые, присутствуют у представителей обоих полов, но их проявление у представителей одного пола сдерживается наличием половых гормонов. Многие физические, различия между полами, вероятно, связаны с признаками этого типа. Поэтому отказ функционирования эндокринных половых желез или нарушение их деятельности может привести к изменению этих признаков. Примером такого ограниченно-полового признака может служить наличие бороды.
Необходимо отметить, что всякий раз, когда какое-либо свойство зависит от единичных признаков, которые определяются единственной генной парой, тогда в результате мы имеем, качественно отличающиеся друг от друга, четко идентифицируемые типы. Однако большинство свойств зависит от множественных факторов, при этом с ростом количества факторов число получающихся в результате комбинаций резко уве-
Наследственность и среда 81
личивается1. Даже когда число влияющих факторов сравнительно невелико, получающиеся в результате индивидуальные различия являются количественными и их распределение может приближаться к нормальному графику. Человеческий рост может быть иллюстрацией такого мультифакторного признака.
Когда речь идет о мультифакторном признаке, проявление или непроявление самого признака может зависеть от единичного фактора. Иными словами, действие множественных факторов зависит от наличия особого гена, который поэтому может рассматриваться как ограничивающее условие. В качестве иллюстрации уместно вспомнить пример с альбиносностью. Сегодня известно, что цвет человеческих глаз зависит от присутствия нескольких генных пар. Из различных комбинаций таких генов возникает почти непрерывный цветовой ряд. Однако если индивид от обоих родителей (ее) получил конкретный фактор, вызывающий альбиносность, то он будет альбиносом, независимо от того, какую комбинацию генов, связанных с цветом глаз, он несет. Влияние всей комбинации генов упраздняется присутствием генной пары альбиносности. Подобно этому, пятнистая шкура у определенной породы крупного рогатого скота возникает от действия единичного рецессивного фактора. Но степень пятнистости образует фактически непрерывный ряд и зависит от множества модифицирующих множественных факторов. Этот тип отношений особенно соответствует возможной роли наследственности в развитии некоторых психологических характеристик. У нас будет случай вернуться к этому при обсуждении определенных типов слабоумия (ср. гл. 12).
И наконец, существует понятие генного баланса. Анализируя материал, генетик обязательно должен изучить влияние конкретных генов на развитие каждого признака. Как бы то ни было, мы должны помнить, что каждый признак в действительности является результатом взаимодействия всех генов, унаследованных ин-
1 В этой связи генетики говорят о главных генах, которые действуют как единичные факторы, производящие эффект распознавания, и полигенах, которые, со своей стороны, влияют на совокупное развитие мультифакторных признаков. Один и тот же ген, конечно, может действовать в качестве главного гена в этиологии одного признака и выступать как полиген в этиологии другого признака.
82 Дифференциальная психология
дивидом. Снайдер и Дэвид (23, с. 251) так выражают современную точку зрения генетиков по этому поводу: «Ген всегда оказывает действие при наличии других генов; отсюда возникает идея генного баланса, согласно которой любой признак является результатом целого генного комплекса, действующего в данной среде. Изменчивость признака может быть следствием изменчивости в единственном гене, но всегда п
|
|
|
