 |


Верхнепермские пресмыкающиеся
|
|
|
|
ПОВОЛЖЬЯ И СЕВЕРНОЙ ДВИНЫ
«Мир прошедший, покорный мощному голосу науки, поднимается из могилы свидетельствовать о переворотах, сопровождавших развитие поверхности земного шара; почва, на которой мы живем, эта надгробная доска жизни миновавшей, становится как бы прозрачною; каменные склепы раскрылись; внутренности скал не спасли хранимого ими. Мало того, что полуистлевшие, полуокаменелые остовы обрастают снова плотью, палеонтология стремится раскрыть закон соотношения между геологическими эпохами и полным органическим населением их. Тогда всё некогда живое воскреснет в человеческом разумении, всё исторгнется от печальной участи бесследного забвения, и то, чего кость истлела, чего феноменальное бытие совершенно изгладилось, восстановится в светлой обители науки, в этой области успокоения и увековечения временного».
Герцен А. «Письма об изучении природы».
Полн. собр. соч., т. IV, стр. 165.
Пермский период — один из наиболее интересных в истории пресмыкающихся и в истории наземной жизни вообще.
Аридизация климата на значительных участках континентов, особенно в верхней перми, и дифференцировка суши сравнительно с предшествовавшим каменноугольным периодом были несомненно весьма важными факторами в истории развития и в характере распространения пермской флоры. За флорой следовала в своем распространении и в своих изменениях и наземная фауна с ее сложными цепями питания и возраставшими в своем разнообразии «нишами жизни».
Что касается самих пресмыкающихся, то из предшествующей каменноугольной эпохи до нас дошли сравнительно немногочисленные остатки этого когда-то обширнейшего класса позвоночных. Пермский же период оставил, как известно, богатейшее наследие. Пресмыкающие достигли в это время, особенно в позднепермскую эпоху, замечательного разнообразия в своих приспособлениях к окружающей среде и в направлениях своей эволюции до млекопитающеподобных включительно. Эта эпоха была, видимо, и временем зарождения бесчисленных пресмыкающихся мезозоя.
|
|
|

Рис. 72. Схема палеогеографии верхней перми для части восточного полушария(по Термье, 1952)
И сами палеонтологические материалы из пермских отложений разных материков, и результаты изучения этих материалов представляют собою в настоящее время итог огромного труда многих поколений ученых разных стран, богатейшее общее достояние единой мировой науки: по своему существу палеонтология, эта «зоология и ботаника угасших форм жизни», как ее называл академик И. П. Павлов, еще более международна, чем зоология современности. Но по специфике палеонтологии, из-за неполноты геологической летописи, по палеозоогеографическому, палеоклиматическому и т. д. разнообразию разных стран, вклад каждой страны в это общее дело, естественно, очень различен. Он часто различен по своему основному исходному материалу в виде остатков животных, но не редко и по выводам на основе изучения этих документов далекого прошлого, а также местонахождений ископаемых остатков.
Не имея возможности подробно останавливаться на истории изучения пермских пресмыкающихся СССР, отметим лишь, что первые упоминания о находках в России относятся еще к XVIII в., изучение же наших пермских рептилий началось в первой половине XIX в. и связано с именами С. Куторги (1838, 1842, 1844), Г. Фишера (1840—1847), Э. Эйхвальда (1861). На рубеже XIX и XX вв. (1895—1903) были поставлены крупные раскопки В. П. Амалицкого (1860—1917) в верхнепермских отложениях р. Северной Двины; итог раскопок — огромные палеонтологические сборы верхнепермских пресмыкающихся, а также земноводных. Эти работы впервые установили близость найденной фауны к южноафриканской из «формации Карру» и послужили научной и организационной основой для последующего систематического изучения палеозойских пресмыкающихся и земноводных. Это стало реальным только после Октябрьской революции, когда огромное плановое развитие всей жизни Советского Союза и его науки, в том числе естествознания, обеспечило возможность быстрого роста и палеонтологии позвоночных. Он связан с именами наших выдающихся ученых — академика П. П. Сушкина (1868—1928) и в особенности основателя и директора Палеонтологического института Академии наук СССР А. А. Борисяка (1872—1944). Непосредственным планированием и выполнением палеонтологических экспедиций для сборов ископаемых палеозойских и мезозойских пресмыкающихся и земноводных, их изучением — лично и силами сотрудников — мы почти целиком обязаны И. А. Ефремову; в его работах (1954 и др.) со-
|
|
|

Профессор В. П. АМАЛИЦКИЙ
(1860—1917)
держится интересный очерк истории исследования низших четвероногих нашей территории. Настоящий очерк имеет целью дать краткий обзор итогов изучения некоторых пермских пресмыкающихся СССР за последнее пол-столетие. При этом имеются в виду пресмыкающиеся верхней перми: именно верхнепермские отложения СССР дали наиболее разнообразный интересный материал. Изучение наших нижнепермских пресмыкающихся находится пока еще в начальной стадии. Обзор всего обнаруженного и изученного за последние годы в СССР вышел бы далеко за пределы настоящего очерка. Имеется в виду рассмотрение лишь двух фаун, точнее «фаунистических комплексов» пресмыкающихся. Одна из них происходит из континентальных отложений, соответствующих верхней части казанского яруса морских отложений и, в общем, по своему характеру и стратиграфическому положению зоне Tapinocephalus серии Бофорт Южной Африки (И. А. Ефремов, 1937—1954). Другая происходит из верхнепермских отложений, относимых к татарскому ярусу (соответствует зоне Cistecephalus Южной Африки) — из слоев, по-видимому, отделенных от содержащих фауну казанского времени перерывом в осадкообразовании.
Остановимся на верхнепермской фауне пресмыкающихся и рассмотрим фауну села Ишеева, около г. Тетюши по среднему течению Волги.
|
|
|
Высокий правый берег Волги около г. Тетюши сложен верхнепермскими песками и глинами с прослоями известняков. Местами все они смяты в причудливые складки, привлекающие внимание пассажиров волжских пароходов, но в основном залегают горизонтально. Западнее, километрах в тридцати, верхнепермские отложения, относимые к казанскому ярусу, выступают в глубоких оврагах, впадающих в живописную долину р. Свияги, которая течет меридионально, как и Волга, но с юга на север.
Здесь, в верхнепермских песках, летом 1929 г. студент-геолог Носов нашел скелет ископаемого позвоночного. Место находки — верховье Каменного Оврага, выходящего в речку Сухую Улему — пересыхающий летом правый приток Свияги, около села Ишеева. Скелет не удалось добыть — хрупкие кости рассыпались. К счастью, находка была сфотографирована местным краеведом Князевым, и этот снимок, опубликованный позднее в журнале «Природа», позволил установить принадлежность скелета древнему хищному ящеру (Б. А. Штылько и А. Н. Рябинин, 1932). В 1930—1931 гг. здесь вел раскопки центральный геологоразведочный институт (ныне ВСЕГЕИ, Ленинград), а в 1934, 1935 и 1939 гг. Палеонтологический институт Академии наук СССР (рис. 73). Эти работы обнаружили остатки интереснейших верхнепермских пресмыкающихся, а

Академик П. П. СУШКИН
(1868—1928)

Рис. 73. Раскопки в Каменном овраге около Ишеева
также земноводных и рыб. Уже первые предварительные описания И. А. Ефремова (1937—1940) были так интересны, что быстро вошли в мировую науку, а скелеты и черепа, украшающие музей Палеонтологического института Академии наук СССР в Москве, могут считаться гордостью этого учреждения.
Раскопки около Ишеева дали главным образом древних терапсид — пресмыкающихся из числа обладавших одной височной впадиной и дугой («подкласс» синапсиды). Для древних терапсид — титанозухий и тапиноцефалов — характерны высокая, как бы вдавленная затылочная сторона черепа, большое отверстие непарного теменного глаза на возвышении теменных костей (рис. 74, 75, 79, 80); нёбо имеет древний тип строения («первичное»).
|
|
|
Для удобства изложения начнем рассмотрение этой фауны с титанозухий, хищных терапсид семейства бритоподид (Brithopodidae Efremov, 1954). Это пресмыкающиеся, напоминающие южноафриканских титанозухий, но более примитивные. Как и улемозавр (см. ниже), они еще сохранили многие черты (рис. 74—77) своих предков, сфенакодонтных пеликозавров, известных из
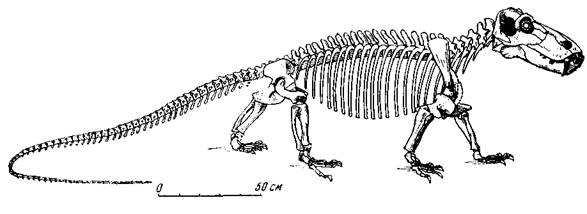
Рис. 74. Титанофонеус (Titanophoneus potens Efremov)
Скелет. Верхняя пермь, Татарская АССР Музей Палеонтологического института АН СССР


Рис. 75. Титанофонеус (Titanopnoneus potens Efremov)
а — череп с нижней челюстью, вид сбоку; б — бугры нёбных костей
с зубами, вид снизу и спереди. Верхняя пермь, Татарская АССР
(Ю. А. Орлов, 1958)
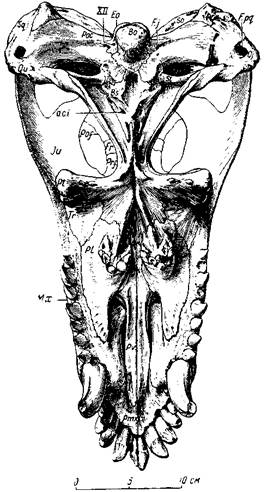
Рис. 76. Титанофонеус (Titanophoneus potens Efremov)
Череп, вид снизу. Верхняя пермь, Татарская АССР
нижнепермских красноцветных толщ Северной Америки.
Наиболее полно изученный титанозух ишеевской фауны — титанофонеус (греческое — «титанический убийца»). В Музее Палеонтологического института хранится почти полный скелет этого пресмыкающегося длиною в три метра (рис. 74). Череп (рис. 75, а) высокий, задняя часть черепа, очень характерная для титанозухий, с отверстием теменного глаза, лежащем на заднем краю черепа на высоком бугре, образованном теменными костями. Нерв теменного глаза был при этом довольно длинным. Затылочная сторона черепа высокая и широкая, что говорит о сильной шейной мускулатуре. Большой срединный вертикальный затылочный гребень указывает на сильную срединную затылочную связку, что отчасти связано с тяжелым черепом (она облегчает своей упругостью приведение головы из опущенного положения в нормальное). Большая височная впадина и сильное развитие гребней на окружающих ее костях, например на теменных, указывают на мощность височной мускулатуры и, вероятно, на стремительное и сильное захлопывание пасти.
Глазные впадины титанозухий направлены косо в стороны и довольно сильно вперед, что для хищника важно при высматривании добычи. Глазное яблоко снабжено кольцом плоских, плотно сочлененных друг с другом косточек, которые увеличивали устойчивость склеры (белочной оболочки глаз). На переднем конце черепа, сжатого с боков, нижний край морды поднимается вперед и вверх, что придает черепу некоторую бульдогообразность; это унаследовано от нижнепермских пеликозавров и, вероятно, связано со способом хватания добычи. Острые резцы направлены косо вперед, особенно верхние, очень плотно входящие в промежутки между нижними (как и нижние между верхними) (рис. 75, а). Точность в смыкании челюстей, без которой животное при стремительном хватании добычи могло бы сломать себе зубы, обеспечена строением челюстного сустава (глубокий двойной блок вверху и двойная впадина в нижней челюсти), возможно, и сильным развитием поперечных отростков крыловидных костей, вероятно, с упором их изнутри в нижнюю челюсть при закрывании рта (рис. 76). Боковые движения нижней челюсти были при этом совершенно исключены. Клыки изогнуты назад и имеют большие корни, глубоко сидящие в челюстях. Задний край клыка снабжен острым режущим гребнем, который мелко зазубрен, как и гребни, отходящие от вершин всех других зубов к их основаниям (такая зазубренность характерна для зубов многих хищных животных, например крупных акул, многих древних пресмыкающихся, ископаемых кошек — «саблезубов» и др.). Послеклыковые, щечные зубы — режущие, но редкие, небольшие, частью отогнуты назад (рис. 75, б; 76). В верхней челюсти для вершин нижних клыков имеются особые углубления.
|
|
|
Своеобразно строение небных костей — каждая снабжена бугром, из которого торчит вниз около десятка небольших цилиндрических зубов с вершиной в виде тупого конуса (рис. 75, б, 76). Массивные поперечные ветви крыловидных костей, примыкающих сзади к небным, внешними утолщенными концами опускаются далеко вниз. На их нижней стороне у титанофонеуса также по два-три зуба, сходных с небными, но посаженных довольно странно — почти горизонтально, вершинами в стороны ветвей нижней челюсти.
Затылочный суставной бугор отогнут вниз, очевидно, голова обычно была наклонена концом морды довольно сильно вниз.
Неясно восприятие звуков титанофонеусом. Столбик уха (stapes, «стремечко») имеет вид небольшой распорки в основании черепа между квадратной костью и мозговой коробкой, где он упирается в овальное окно. Это положение столбика как будто бы исключает его подвижность и приспособленность для передачи звуковых колебаний через овальное окно во внутреннее ухо, хотя по местоположению такой столбик равнозначен стремени среднего уха млекопитающих, птиц и пресмыкающихся, имеющих барабанную перепонку и воспринимающих звук. Как воспринимали звук титанофонеус и другие пресмыкающиеся с таким строением — неясно, но вряд ли они были глухие. Впрочем, змеи по-своему «слышат», хотя лишены барабанной перепонки, и «столбик уха» у них, прежде всего — механическая конструктивная часть черепа, а не слуховая (см. ниже).
Позвоночный столб титанофонеуса сравнительно с черепом стройный и легкий. В нем около сотни двояковогнутых позвонков с тонким отверстием для хорды. Шейный позвонок, следующий за атлантом (эпистрофей), снабжен массивным остистым отростком. Это, вероятно, также связано с большим весом тяжелой головы, что требовало сильной затылочной связки и мускулатуры. Тела позвонков на переднем и заднем концах с округлыми внешними краями, что указывает на гибкость животного и его подвижность. Три крестцовых позвонка соединены неподвижно с тазом крестцовыми ребрами. Хвост длинный, в нем около 60 позвонков из всех ста. Подвздошная кость по строению промежуточна между таковой древнейших пресмыкающихся, где она вытянута назад, и подвздошной более поздних млекопитающеподобных рептилий, у которых верхний отдел кости вытянут вперед, как у млекопитающих.
Передние и задние конечности титанофонеуса одинаковой длины, сильные, стройные, хорошо подвижные; плечо с предплечьем, бедро и голень очень напоминают крокодильи; при этом лапы относительно роста животного очень широкие, главным образом за счет пястных и плюсневых костей, увеличивающихся от первого пальца к пятому (рис. 77). Большая величина лап особенно заметна, если сравнить титанофонеуса с крокодилом, даже вдвое или втрое большим, чем титанофонеус. У титанофонеуса при наличии перепонки между пальцами — это «почти ласты», скорее всего предназначенные для плавания, а также передвижения по влажному и вязкому, илистому побережью рек и озер. Крокодилы обычно плывут вперед, работая высоким хвостом, сжатым с боков наподобие весла, а также боковыми изгибами тела. Длинным хвостом титанофонеус, вероятно, мог наносить сильные удары, как это делают крупные ящерицы и крокодилы, но для плавания хвост не был приспособлен. Титанофонеусу могли давать быстроту в воде его расширенные на концах передние и задние лапы, а также боковые изгибы тела. Число фаланг в пальцах на первый взгляд то же, что и у многих млекопитающеподобных пресмыкающихся и млекопитающих, т. е. та же «фаланговая формула» — 2. 3. 3. 3. 3, от первого пальца к пятому. Но вторая фаланга четвертого пальца передней лапы срослась из двух: из них одна, сочленяющаяся с первой основной фалангой пальца, совсем короткая, а следующая, сочлененная с ногтевой, сохранила длину, близкую к нормальной (рис. 77). Очевидно, четвертый палец близких предков титанофонеуса еще «недавно» обладал четырьмя самостоятельными фалангами.
Титанофонеус явно хищник, но вряд ли часто охотился на своих крупных сородичей, растительноядных тапиноцефалов, улемозавров (рис. 80—82) и вообще на такую крупную добычу. Верхние резцы титанофонеуса, отчасти и нижние, направлены косо вперед; верхний клык, хотя изогнут назад, но не так велик, чтобы мог считаться предназначенным для разрывания на части взрослых улемозавров, величиной с самого титанофонеуса, или крупнее его. Сравнительно слабые послеклыковые зубы, хотя и режущие, но редкие, мало удобны для измельчения мышц, связок и грубой кожи крупных пресмыкающихся, а, скорее всего, приспособлены для удержания добычи. Из всех зубов наиболее мощные — клыки и предклыковые зубы, видимо, по преимуществу, хватательные, хотя гребни на краях верхних резцов, заходящие за края нижних, делали резцы одновременно и режущими. Небольшой уступ на внутренней, язычной стороне резца, в который упиралась вершина зуба противоположной челюсти — «пятка», делал резцы отчасти измельчающими пищу, однако у титанофонеуса лишь в слабой степени. Наиболее совершенны в этом отношении резцы улемозавров (рис. 81) (И. А. Ефремов, 1940).
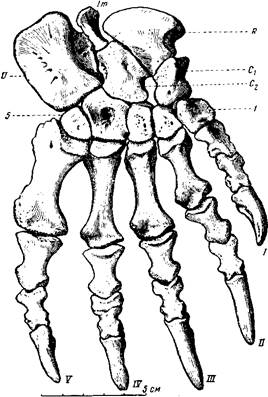
Рис. 77. Титанофонеус (Titanophoneus potens Efremov)
Правая кисть: U — ulnare; Im — intermedium; R — radiale;
C1C2 — carpalia. Верхняя пермь, Татарская АССР (Ю. А. Орлов, 1958)
Небные зубы (рис. 75, б), торчащие вершиной вниз из бугров небных костей, при отсутствии противопоставленных им внизу, не приспособлены для жевания. Вероятно, мы имеем дело с каким-то своеобразным раздавливающим, а частично и удерживающим добычу приспособлением. Для раздавливания добычи могли служить отчасти и поперечные массивные отростки птеригоидов.
Если принять во внимание и «крокодилье» строение плеча с предплечьем, бедра с голенью и широкие лапы титанофонеуса, то можно прийти к предположению о приспособленности животного к полуводному образу жизни и питанию рыбой с плотной чешуей (остатки рыб в этом местонахождении довольно часты), моллюсками с тонкой раковиной, мелкими земноводными и пресмыкающимися, а также другой некрупной добычей. Титанофонеус мог ее ловить и, высматривая с берега, что было удобно при обычно наклонном положении головы, мордой книзу (рис. 74), и в воде, так как он, вероятно, был хорошим пловцом. Разумеется, титанофонеусы, собравшись вместе, были небезопасны и для улемозавров (см. ниже), кормившихся растительностью на побережье верхнепермских озер и рек, и могли стащить в воду и разорвать зазевавшегося улемозавра, но по своему образу жизни вряд ли были повседневными врагами крупных улемозавров, вообще крупной добычи.
Другой, близкий к титанфонеусу, но более грозный титанозух из раскопок в Каменном Овраге — хищный долиозавр (греческое долиозавр — коварный ящер) (рис. 78), принятый вначале за большого старого самца титанофонеуса. Известно, что многие крупные низшие позвоночные (крокодилы, черепахи, щуки и др.) очень долговечны, а главное обладают длительным ростом уже во взрослом, половозрелом состоянии. Поэтому разница между долиозавром и титанофонеусом в размерах сначала не казалась сама по себе существенной. Позднее выяснялись отличия в строении черепа, зубов и т. д., и «старый самец титанофонеуса» оказался новым очень крупным хищным титанозухом.
Тяжелый, высокий череп долиозавра достигает в длину более полуметра. Многие кости верхней стороны черепа очень утолщены (особенно лобные, окружающие глазницу, и теменные); снаружи они сильно бугристы с бороздами кровеносных сосудов и отдаленно напоминают бугристость костей, несущих рога, например у носорогов из млекопитающих. Вероятно, у долиозавра это указывает на очень сильное ороговение кожи защитного характера. Морда, сжатая с боков, высокая; глазничные впадины, окруженные массивными костями, невелики. Резцы и особенно клыки крупные, в основном приспособленные для хватания добычи при стремительном захлопывании пасти, на которое указывают большие височные впадины, вмещавшие челюстную мускулатуру. Послеклыковые зубы слегка копьевидны, малы, не образуют сомкнутого ряда, немногочисленны и скорее предназначены для удержания добычи, чем для ее измельчения. Небные бугры не-
Рисунок на вкладке (файл orl132r.jpg: Титанофонеусы и улемозавры. Рис. К. К. Флерова)
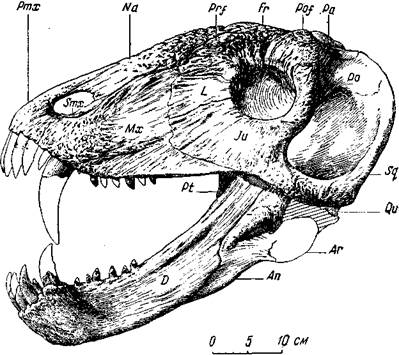
Рис. 78. Долиозавр (Doliosaurus yanshinovi Orlov)
Череп, вид сбоку. Верхняя пермь. Татарская АССР (Ю. А. Орлов, 1958)
велики, с мелкими тупоконическими зубами: позади небных бугров расположено по два-три таких же зуба на птеригоидах; зубы на поперечных отростках птеригоидов мелкие. Помимо впадин, для нижних клыков (впереди верхних), что характерно для многих хищных титанозухов и родственных им пресмыкающихся, на небе долиозавра имеются ямки и для нескольких послеклыковых, «щечных» зубов. Их вершины должны были входить в эти небные ямки довольно точно во избежание поломки, а объединенность ямок в одном общем желобообразном углублении, быть может, указывает на возможность небольших, по только передне-задних движений нижней челюсти; об этом же говорит и несколько уплощенная форма челюстного сустава. При захлопывании пасти нижние послеклыковые зубы ложились внутрь от верхних, не соприкасаясь с ними. Это подтверждает предположение о приспособленности щечных зубов для удержания добычи, а не ее измельчения.
Не совсем понятна необычайная (для размеров животного) массивность лобной и теменной частей черепа. Частично это можно было бы толковать как итог приспособления к большой механической нагрузке со стороны переднего конца морды при хватании и удержании крупной добычи. Однако это не объясняет странного бугристого строения внешней поверхности костей верхней стороны черепа. (Существует предположение, что большой толщины кости крышки черепа служили и защитой от перегрева в жаркое время, но это допущение не особенно убедительно). Остальной скелет в общем похож на скелет титанофонеуса, но крупнее и гораздо массивнее. Долиозавр был явно хищником, но характер его пищи не совсем ясен. Более массивный и, видимо, менее подвижный, чем титанофонеус, долиозавр вряд ли был особенно проворным, активным охотником за рыбой и другими водными животными, которых, правда, мог и подстерегать. Описываемая фауна была богата, кроме всего прочего, небольшими мелкими земноводными и пресмыкающимися; для них в то время долиозавр, вероятно, был поистине «царем зверей». Впрочем, известно, что хищные пресмыкающиеся, даже крупные, например огромные удавы и крокодилы, с их низкой, переменной температурой крови гораздо менее прожорливы, чем это может показаться на первый взгляд.
Небольшой хищный титанозух сиодон (рис. 79) с черепом всего 20 см длины, казалось бы, очень походит на титанофонеуса. Но теменной глаз у сиодона относительно крупнее; его отверстие лежит на тонкостенном бугре, относительно более высоком, чем у титанофонеуса. Верхние клыки сиодона изогнуты назад сильнее, в поперечнике округлые, без режущего заднего края, без острой колющей вершины. Послеклыковых зубов много, они низкие, тупоконические, менее сжатые с боков, стоят прямо, часто, в отличие от зубов титанофонеуса, хорошо приспособлены для измельчения пищи. Небные бугры с зубами, как и поперечные отростки птеригоидов, относительно сильнее, чем у титанофонеуса. Но особенно резко отличны все послеклыковые челюстные зубы. Вдобавок задние из них иногда стоят в нижней челюсти не только друг за другом, но и рядом, попарно. Такая «пара» однобугорчатых зубов — своего рода заменитель, хотя и не очень совершенный, одного, но бугорчатого зуба некоторых пресмыкающихся (более поздних, триасовых), а также млекопитающих. По-видимому, сиодон мог быть всеядным животным, хотя в основном, вероятно, охотился за разными мелкими животными. Большие глазницы наводят на предположение о значительной величине глаз и о сумеречном или ночном образе жизни этого животного.
Добыты остатки более дюжины особей сиодона, но, к сожалению, ни одного цельного скелета; видимо, этому животному принадлежат многочисленные кости конечностей и позвонки не-

Рис. 79. Сиодон (Syodon efremovi Orlov)
Череп, вид сбоку. Верхняя пермь, Татарская АССР
(Ю. А. Орлов, 1958)
большого титанозуха ишеевской фауны. По костям конечностей и позвонкам сиодон походил на титанофонеуса, но был много меньше его (около полутора метров в длину). Лопатка у титанофонеуса и у сиодона изогнута очень слабо; между тем ее кривизна при близком расположении лопатки к грудной клетке в известной степени отражает форму клетки: вполне возможно приписать титанофонеусу и сиодону сжатое с боков туловище, а не уплощенное сверху вниз, как обычно у крокодилов.
Крупное растительноядное пресмыкающееся этой фауны — тапиноцефал улемозавр (рис. 80) [5]. Череп улемозавра имеет очень высокую, наклонную назад затылочную сторону; верхний край глазницы так сильно утолщен, что сбоку кажется лобным рогом; лобные и все окологлазничные кости бугристы на внешней поверхности и так массивны, что при рассматривании черепа сверху совсем не видно глазных впадин: они прикрыты «нахмуренным» лбом и задними краями глазниц. При длине всего черепа около 40 см с небольшим толщина теменных костей не
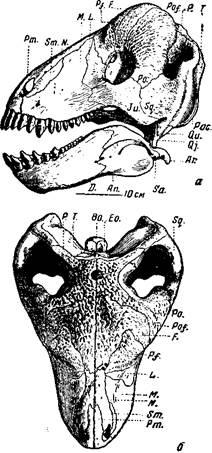
Рис. 80. Улемозавр (Ulemosaurus svijagensis Rjabinin)
а — череп, вид сбоку: б — вид сверху. Верхняя пермь, Татарская АССР
(И. А. Ефремов, 1940)
менее 7 см (очевидно, еще больше была длина нерва теменного глаза). Утолщение теменных, скуловой, всех окологлазничных костей и костей височной области сужает височную впадину, вмещавшую височный мускул (особенно в ее нижней части) в относительно узкую щель.
Затылок пропорционально выше, чем у титанозухий, поперечные отростки крыловидных костей развиты слабее, не доходят до ветвей нижней челюсти и не ограничивают ее движений; челюстной сустав допускает некоторую свободу боковых движений, что обычно характерно для
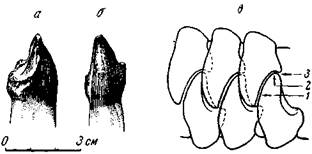
Рис. 81. Улемозавр (Ulemosaurus svijagensis Rjabinin)
а — нижний третий правый резец улемозавра, вид сбоку; б —
спереди; в — схема режущей, перетирающей и раздавливающей
работы резцов улемозавра. Верхняя пермь, Татарская АССР
(И. А. Ефремов, 1940)
растительноядных (и всеядных) позвоночных. Скуловая дуга, морда и нижняя челюсть короче, выше и массивнее, чем у хищных дейноцефалов. Нёбо сходно с нёбом титанозухий, но улемозавр лишен бугров с зубами на нёбных костях и без зубов на крыловидных. Столбик уха, «стремечко», как и у титанофонеуса, имеет вид распорки и в основании черепа между квадратной костью, сочлененной с нижней челюстью, и мозговой коробкой. Столбик закрывает своим расширенным концом овальное окно, ведущее в лабиринт внутреннего уха; столбик еще массивнее, чем у титанофонеуса, и на первый взгляда еще менее может быть заподозрен в способности проводить звук; в общем, восприятие звука здесь столь же неясно, как и у титанофонеуса. У некоторых современных пресмыкающихся, по своему образу жизни «прижатых» к земле (например, змей и некоторых ящериц), звуковые колебания, по-видимому, передаются внутреннему уху через кости черепа; однако такой образ жизни трудно предположить для титанофонеуса, а особенно для улемозавра, голова которого, судя по строению плечевого пояса, передних ног и позвоночного столба, была обычно довольно высоко поднята над землей (рис. 82).
Отогнутый вниз затылочный сочленовный бугор черепа, нависший над ним затылок и форма скошенных на переднем и заднем концах шейных позвонков указывают на наклонное положение головы улемозавра (концом морды обычно вниз).
Зубы улемозавра, около 15 на каждой стороне вверху и внизу, необычного для пресмыкающихся вида, но типичного для растительноядных тапиноцефалов. Самые интересные — передние, «резцы» (рис. 81). Они много крупнее остальных, в связи с этим передние концы межчелюстных и нижнечелюстных костей очень массивны. Далее назад зубы постепенно уменьшаются (рис. 80, а); исключение составляют снабженные режущими краями клыки, хотя они почти утратили клыкообразность. В основании коронки резцов верхних и нижних, на их язычной стороне, очень сильно развит выступ, так называемая «пятка»; в ее углубление входит вершина резца — антагониста противоположной челюсти. Эмаль зубов по краям коронок острая, зубчатая. При смыкании челюстей вследствие частичного вклинивания нижних зубов в промежутки между верхними (и наоборот) резцы одновременно перерезают, раздавливают и перетирают пищу (рис. 81, в), чему и должен был соответствовать характер работы челюстной мускулатуры. Таким образом, эти зубы — резцы по местоположению — одновременно выполняют и работу коренных зубов, будучи хорошо приспособлены для измельчения пищи, вероятно, каких-то мясистых и сочных растений. Послеклыковые зубы мелкие с острыми режущими передними и задними краями (как у относительно слабого клыка) и, скорее всего, удобны для перекусывания растений.
Для тапиноцефалов (рис. 82) — растительноядных пресмыкающихся, к которым относятся улемозавры, характерны очень сильные высокие передние ноги, массивный передний пояс (лопатка, коракоиды, ключицы, межключица). Спина своеобразно поката назад, на лапах последние фаланги в виде мелких копытец; сильные передние конечности, возможно, служили и для добывания пищи и, видимо, вообще выполняли наибольшую работу, задние относительно слабые. Строение конечностей, плечевого пояса и таза указывает на возможность высоко приподнятого над землей положения животного, подобно млекопитающим (как это, впрочем, иногда наблюдается и у передвигающегося по суше крокодила). Для улемозавра такое приподнятое над землей положение было, скорее всего, обычно. Вряд ли были особенно проворны улемозавр и его ближайшие родичи, другие тапиноцефалы, более древние — дейтерозавры из верхнепермских отложений Южного Приуралья и тем более — поздние и громоздкие южноафриканские мосхопс, мормозавр и др. Но все тапиноцефалы были, очевидно, хорошо приспособлены к растительной пище и, вероятно, поэтому распространились так широко. Это первые, наиболее древние известные крупные раститель-
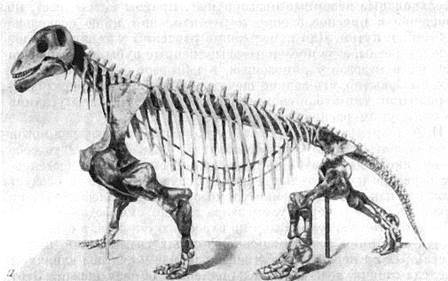

Рис. 82. Мосхопс (Moschops capensis Broom), близкий к улемозавру
растительноядный дейноцефал из верхнепермских отложений формации
Карроо Южной Африки
а — скелет; б — реконструкция внешнего вида (W. Gregory, 1926)
ноядные пресмыкающиеся, вообще первые крупные растительноядные наземные позвоночные, притом «копытные», но пошедшие в приспособлении к растительной пище несколько необычным путем. Для измельчения растений у тапиноцефалов служили резцы, а не послеклыковые щечные зубы, как это было позднее, в мезозое у динозавров, в кайнозое — у млекопитающих. Любопытно, что задние щечные зубы улемозавра сохранили режущий тип зубов своих предков, пожалуй, даже усилили его сомкнутым расположением зубов.
И. А. Ефремов (1940) показал, что резцы улемозавров могли легко развиться из зубов титанозухий, если представить себе увеличение «пятки» зуба на его язычной стороне при постепенном переходе к растительной пище. Разумеется, это не следует понимать как происхождение улемозавра от титанофонеуса: не только зубы, но и череп улемозавра можно «вывести» от титанофонеуса, так как улемозавр по строению скелета, в сущности говоря, есть лишь видоизменение хищных титанозухий, преобразованных на первый взгляд до неузнаваемости под влиянием перехода хищника к растительноядному образу жизни. Этот переход вызвал перемену нагрузок на зубы, челюсти, череп и потребовал значительной его перестройки. Но улемозавр и титанофонеус — современники, связанные общим происхождением от еще более древних и архаичных по строению терапсид, предками которых были сфенакодонтные пеликозавры, разнообразные в нижнепермских отложениях Северной Америки.
Не менее замечательно и другое загадочное пресмыкающееся ишеевской фауны — венюковия (рис. 83). Родовое название было дано В. П. Амалицким в честь П. Н. Венюкова (1858—1915), профессора Петроградского горного института.
Первая находка, четыре обломка нижней челюсти, происходит из верхнепермских медистых песчаников Южного Приуралья, близких по геологическому возрасту к пескам Ишеева, но, вероятно, более ранних. Эти остатки были собраны П. Н. Венюковым в отвалах старых медеплавильных рудников в районе Оренбурга и переданы в 1908 г. профессору В. П. Амалицкому. Торчащие вперед сточенные нижние резцы венюковии очень похожи на долотообразные резцы грызунов. Это сходство, несомненно, и было причиной ошибки В. П. Амалицкого (1922*), который отнес животное к млекопитающим. Истинная природа венюковии была выяснена И. А. Ефремовым лишь после раскопок около Ишеева в 1935 г., когда стали попадаться такие же обломки нижних челюстей, а затем нашелся и череп.
Венюковия из описываемой фауны поражает массивностью конца морды, особенно нижней челюсти (рис. 83). Межчелюстные кости необычайно развиты и утолщены; на нёбе они дохо-
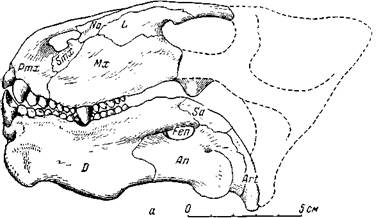

Рис. 83. Венюковия (Venjukovia invisa Efremov)
а — череп, вид сбоку (И. А. Ефремов, 1940); б — нижняя челюсть с куском левой верхней челюсти, вид сверху (Ватсон, 1948). Задние концы правой и левой ветвей нижней челюсти должны быть раздвинуты шире, судя по другим образцам (сжаты деформацией после захоронения). Верхняя пермь, Татарская АССР
дят до нёбных, а снаружи черепа почти до уровня глазниц. Сильно утолщены и верхнечелюстные кости; а высота нижней челюсти та же, что самого черепа впереди глазниц. Особенно развита передняя часть нижней челюсти; ее передний край сильно утолщен, внизу справа и слева имеются массивные выступы, спереди придающие симфизу вид раздвоенного подбородка, опускающиеся значительно ниже симфиза. Пожалуй, самое замечательное у этого пресмыкающегося — его зубы, хотя сами по себе они имеют простое строение.
Самые крупные — резцы верхней челюсти, второй и третий, низкие, толстые; слегка вогнутые площадки стирания на коронке сходятся под углом на вершине зуба. Далее назад следует после нескольких мелких конических зубов крупный, низкий, толстый клык со слабо стирающейся коронкой. Он входит при смыкании рта своей вершиной в особую ямку в нижней челюсти снаружи от нижних зубов (рис. 83). Из нескольких мелких конических зубов, следующих в верхней челюсти за клыком, самые последние стоят попарно рядом или почти рядом, а не друг за другом, как обычно. Еще отчетливее это необычное расположение наблюдается в заднем отделе нижнего зубного ряда (рис. 83, б). На переднем конце нижней челюсти — два крупных долотообразных резца. И если передние зубы верхней и нижней челюсти сходны с резцами грызуна, то задние, расположенные в два ряда, по своему назначению, очевидно, соответствуют бугорчатым коренным зубам млекопитающего. Таким образом, венюковия своими зубами напоминает грызунов с их долотообразными резцами и с перетирающими растительную пищу коренными. Однако это сходство с грызунами относительное.
У венюковии не сохранилось заднего отдела черепа, но, судя по уцелевшей глазничной части, задняя половина черепа вряд ли была массивной. Во всяком случае, лицевая часть сравнительно с глазничной очень мощная; особенно же непомерно массивна передняя половина нижней челюсти (рис. 83, а), а ее задняя половина и суставной отдел относительно слабые; послеклыковые зубы мелкие. Если венюковия и обладала передними грызущими зубами, то ее послеклыковые зубы не годятся для перетирания твердой растительной пищи, зерен и т. п., и животное вряд ли питалось, как грызуны. Поэтому И. А. Ефремов (1940) предположил, что само питательное вещество было мягким, но извлекалось из каких-то твердых и крупных оболочек, которые не могли быть непосредственно раздроблены во рту, например раковин, и что венюковия могла питаться крупными растениями с твердой оболочкой вроде современных саговых пальм или кактусов. У некоторых современных грызунов имеются подбородочные выступы на нижней челюсти, как у венюковии. Эти выступы и небольшой гребень на черепе впереди глазницы побудили И. А. Ефремова предположить у венюковии большую силу челюстной мускулатуры.
Мы не знаем современных или ископаемых пресмыкающихся в точности с таким строением нижней челюсти, как у
|
|
|
