 |


Таутомерия и некоторые другие физико-химические свойства оснований
|
|
|
|
Перечисленные пуриновые и пиримидиновые основания содержат сопряженную систему кратных связей и заместители (группы —ОН и —NH2). Указанные структурные особенности обусловливают способность пуриновых и пиримидиновых оснований к различным типам таутомерных превращений: лактам-лактимному для оксипроизводных и амин-иминному для аминопроиз-водных. На примере урацила таутомерные превращения урацила можно представить в следующем виде:
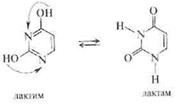
В случае дизамещенных пуринов и пиримидинов теоретическое количество изомерных форм увеличивается, однако практически существуют лишь некоторые из них. По данным ИК- и ЯМР-спектроскопии показано, что в нуклеиновых кислотах пуриновые и пиримидиновые основания преимущественно находятся в лактамной и аминной формах; это обеспечивает правильность спаривания нуклеотидов в ходе матричных синтезов нуклеиновых кислот. Однако под влиянием внешних факторов, например воздействия излучений, возможен переход оснований в другие таутомерные формы, лежащий в основе мутагенеза.
Рентгеноструктурный анализ трехмерной структуры различных пуриновых и пиримидиновых оснований показал, что молекулы пиримидинов имеют абсолютно плоское строение, а молекулы пуринов — почти плоское.
Азотистые основания поглощают свет в ультрафиолетовой области спектра с максимумом около 260 нм. Поглощение в ультрафиолетовой области используется для количественного определения нуклеиновых кислот.
ДНК и РНК, различия между ними. 14.2.3. Углеводные компоненты
Углеводная часть нуклеотидов, входящих в РНК, представлена рибозой, а входящих в ДНК, — дезоксирибозой. Пентозы в нуклеиновых кислотах всегда присутствуют в p-D-фуранозной форме:
|
|
|
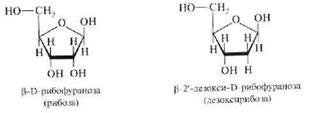
Углеродные атомы пентоз в нуклеотидах нумеруются со знаком «штрих», чтобы их можно было отличить от атомов азотистых оснований.
Доказано, что замена у дезоксирибозы при С-2' группы ОН на протон упрочняет связь между С-2' и С-3'. Это, в свою очередь, увеличивает прочность молекулы ДНК и способствует компактности ее пространственной структуры.
 Локализация ДНК в клетке.1) в прокариотической – цитоплазма;
Локализация ДНК в клетке.1) в прокариотической – цитоплазма;
2) в эукариотической – ядро, органеллы (митохондрии, пластиды, клеточный центр).
Форма молекул ДНК. ДНК, подобно белкам, имеет первичную, вторичную и третичную структуры.
Первичная структура ДНК. Данная структура определяет закодированную в ней информацию, представляя собой последовательность чередования дезоксирибонуклеотидов в полинуклеотидной цепи. Хотя ДНК содержит всего четыре типа мономерных звеньев, количество возможных нуклеотид- Вторичная структура ДНК. В соответствии с моделью, предложенной в 1953 г. Дж. Уотсоном и Ф. Криком, она представляет собой двухцепочечную правозакрученную спираль из комплементарных друг другу антипараллельных полинуклеотидных нитей.
Для вторичной структуры ДНК решающим являются две особенности строения азотистых оснований нуклеотидов. Первая заключается в наличии групп, способных образовывать водородные связи. Так, между А и Т могут образовываться две, а между Г и Ц — три водородные связи. Эти азотистые основания называются комплементарными. Вторая особенность заключается в том, что пары комплементарных оснований А—Т и Г—Ц оказываются одинаковыми не только по размеру, но и по форме (рис.14.2).
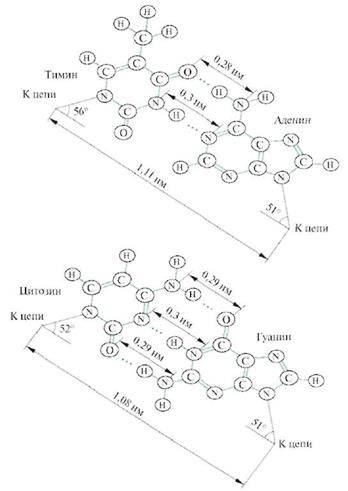
Благодаря способности нуклеотидов к спариванию, образуется жесткая, хорошо стабилизированная двухцепочечная структура, обладающая следующими свойствами (рис. 14.3).
Сахарофосфатные остовы двух цепей образуют правозакрученную спираль с общей осью и диаметром 0,2 нм. В спирали существуют две бороздки — большая и малая. На каждый виток спирали приходится 10 пар оснований.
|
|
|
• Сахарофосфатные остовы двух полинуклеотидных цепей, расположенные снаружи, связаны между собой водородными связями между отходящими от них вовнутрь азотистыми основаниями. Плоскости оснований перпендикулярны оси спирали и отстоят друг от друга на 0,34 нм.
• Гидрофобные взаимодействия между плоскостями ароматических колец оснований стабилизируют структуру, преодолевая силы электростатического отталкивания между отрицательно заряженными фосфатными группами.
• Две цепи антипараллельны, т. е. по своему химическому строению они ориентированы в противоположных направлениях. Антипараллельная направленность имеет важное биологическое значение при репликации и транскрипции ДНК.
На основе тщательного анализа рентгенограмм выделенных ДНК установлено, что двойная спираль ДНК может существовать в виде нескольких форм (А, В, С, Z и др.). Указанные формы ДНК различаются диаметром и шагом спирали, числом пар оснований в витке, углом наклона плоскости оснований по отношению к оси молекулы (рис. 14.4).
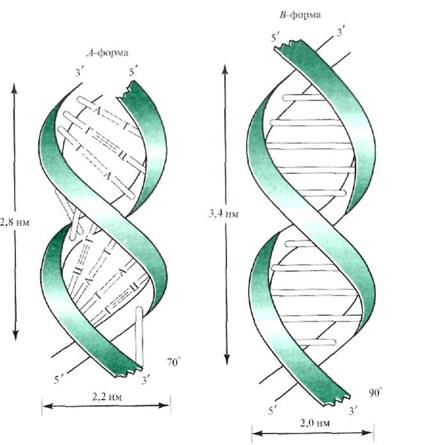
Рис. 14.4. Схема А- и В-форм двойной спирали
Третичная структура ДНК. У всех живых организмов двухспиральные молекулы ДНК плотно упакованы с образованием сложных трехмерных структур.
Двухцепочечные ДНК прокариот, имеющие кольцевую ковалентно-зам-кнутую форму, образуют левые (-) суперспирали. Суперспирализация прежде всего необходима для «упаковки» громадной молекулы ДНК в малом объеме клетки. Например, ДНК Е. coli имеет длину более 1 мм, в то время как длина клетки не превышает 5 мкм. Помимо этого, суперспирализация ДНК, облегчающая ее расплетение, обеспечивает начало репликации и транскрипции (рис. 14.5).
Третичная структура ДНК эукариотических клеток также образуется путем суперспирализации, но не свободной ДНК, а ее комплексов с белками хромосом.
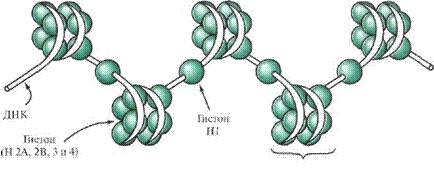
Ядерный хроматин содержит ДНК, гистоновые и негистоновые белки, небольшое количество РНК. В пространственной организации хромосом можно выделить несколько уровней. Первый уровень — нуклеосомный. Нуклеосом-ная нить образуется при взаимодействии ДНК с белками-гистонами. Гистоны представляют собой простые белки с молекулярной массой 14—20 kDa, в аминокислотном составе которых преобладают аргинин и лизин, глицин и цисте-ин. Преобладание лизина и аргинина придает гистонам щелочной характер и обеспечивает их способность взаимодействовать с кислотными группами ДНК. Во всех типах эукариотических клеток обнаружено 5 классов гистонов (HI, Н2, НЗ, Н4, Н5), различающихся по содержанию (%) основных аминокислот, обусловливающему их физико-химические свойства (электрофоретическую подвижность, ИЭТ и др.). Гистоны являются эволюционно консервативными белками. Степень гомологии аминокислотных последовательностей гистонов Н2, НЗ, Н4, Н5 у разных видов животных, растений и грибов достаточно высока. Эти гистоны попарно образуют октамеры (белковые коры дисковидной формы, которые оплетаются молекулой ДНК). Участок ДНК, спирально оплетающий октаметр, содержит в среднем 145—150 нуклеотидных пар и формирует примерно 1,75 витка левой спирали. Свободные от контакта с белковыми корами участки ДНК называют линкерными (или связующими). Их длина варьирует в за-
|
|
|
висимости от типа клеток (от 15 до 100 нм). Линкерные участки ДНК либо свободны, либо связаны с гистоном HI, который способствует компактизации нуклеосомной нити и может препятствовать транскрипции ряда генов. Гистоны в клетках подвергаются ковалентной модификации путем фосфорилирова-ния, ацетилирования, метилирования и др. Это приводит к изменению их способности взаимодействовать с ДНК, что является одним из механизмов регуляции транскрипции генов.
В результате нуклеосомной организации хроматина двойная спираль ДНК диаметром 2 нм приобретает диаметр 10—11 нм и укорачивается примерно в 7 раз.
Вторым уровнем пространственной организации хромосом является образование из нуклеосомной нити хроматиновой фибриллы диаметром 20— 30 нм, что обеспечивает уменьшение линейных размеров ДНК еще в 6—7 раз. Наиболее вероятной считается соленоидная модель упаковки в хроматиновой фибрилле.
|
|
|
Третичный уровень организации хромосом обусловлен укладкой хроматиновой фибриллы в петли. В образовании петель принимают участие негистоновые белки, узнающие специфические нуклеотидные последовательности в ненуклеосомной ДНК и фиксирующие образование петель. Участок ДНК, соответствующий одной петле, содержит от 20 000 до 80 000 пар нуклеотидов и, вероятно, представляет домен ДНК, соответствующий единице транскрипции. В результате такой упаковки линейные размеры ДНК уменьшаются примерно в 200 раз. Петлеобразная доменная организация ДНК, называемая интерфазной хромонемой, может подвергаться дальнейшей компактизации, степень которой меняется в зависимости от фазы клеточного цикла (рис. 14.6).
Типы РНК, их роль в жизнедеятельности клетки. Матричная (информационная) РНК — РНК, которая служит посредником при передаче информации, закодированной в ДНК к рибосомам, молекулярным машинам, синтезирующим белки живого организма. Кодирующая последовательность мРНК определяет последовательность аминокислот полипептидной цепи белка. Однако подавляющее большинство РНК не кодируют белок. Эти некодирующие РНК могут транскрибироваться с отдельных генов (например, рибосомальные РНК) или быть производными интронов. Классические, хорошо изученные типы некодирующих РНК — это транспортные РНК (тРНК) и рРНК, которые участвуют в процессе трансляции. Существуют также классы РНК, ответственные за регуляцию генов, процессинг мРНК и другие роли. Кроме того, есть и молекулы некодирующих РНК, способные катализировать химические реакции, такие, как разрезание и лигирование молекул РНК. По аналогии с белками, способными катализировать химические реакции — энзимами (ферментами), каталитические молекулы РНК называются рибозимами. Участвующие в трансляции (мРНК, тРНК, рРНК, тмРНК). Информация о последовательности аминокислот белка содержится в мРНК. Три последовательных нуклеотида (кодон) соответствуют одной аминокислоте. В эукариотических клетках транскирибированный предшественник мРНК или пре-мРНК процессируется с образованием зрелой мРНК. Процессинг включает удаление некодирующих белок последовательностей (интронов). После этого мРНК экспортируется из ядра в цитоплазму, где к ней присоединяются рибосомы, транслирующие мРНК с помощью соединённых с аминокислотами тРНК.
В безъядерных клетках (бактерии и археи) рибосомы могут присоединяться к мРНК сразу после транскрипции участка РНК. И у эукариот, и у прокариот цикл жизни мРНК завершается её контролируемым разрушением ферментами рибонуклеазами.
Транспортные (тРНК) — малые, состоящие из приблизительно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты в место синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодонам мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединённой к тРНК.
|
|
|
Рибосомальные РНК (рРНК) — каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырёх типов рРНК синтезируются в ядрышке. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеин, называемый рибосомой. Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80 % РНК, обнаруживаемой в цитоплазме эукариотической клетки.
Необычный тип РНК, который действует в качестве тРНК и мРНК (тмРНК) обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию.
РНК-интерференция. В живых клетках обнаружено несколько типов РНК, которые могут уменьшать степень выражения гена при комплементарности мРНК или самому гену. Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и оказывают воздействие через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводить к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградируется. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов. Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам. У животных найдены так называемыме РНК, взаимодействующие с Piwi (piРНК, 29-30 нуклеотидов), действующие в половых клетках против транспозиции и играющие роль в образовании гамет. Кроме того, piRNA могут эпигенетически наследоваться по материнской линии, передавая потомству своё свойство ингибировать экспрессию транспозонов.
Антисмысловые РНК широко распространены у бактерий, многие из них подавляют выражение генов, но некоторые активируют экспрессию. Действуют антисмысловые РНК, присоединяясь к мРНК, что приводит к образованию двуцепочечных молекул РНК, которые деградируются ферментами. У эукариот обнаружены высокомолекулярные, мРНК-подобные молекулы РНК. Эти молекулы также регулируют выражение генов. В качестве примера можно привести Xist, присоединяющуюся и инактивирующую одну из двух Х-хромосом у самок млекопитающих.
Кроме роли отдельных молекул в регуляции генов, регуляторные элементы могут формироваться в 5' и 3' нетранслируемых участках мРНК. Эти элементы могут действовать самостоятельно, предотвращая инициацию трансляции, либо присоединять белки, например, ферритин или малые молекулы, например, биотин.
В процессинге РНК. Многие РНК принимают участие в модификации других РНК. Интроны вырезаются из пре-мРНК сплайсосомами, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК). Кроме того, интроны могут катализировать собственное вырезание. Синтезированная в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышке и тельцах Кахаля. После ассоциации мяРНК с ферментами, мяРНК связываются с РНК-мишенью путём образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК. Гидовые РНК осуществляют процесс редактирования РНК в кинетопласте — особом участке митохондрии протистов-кинетопластид (например, трипаносом).
6. Клеточный цикл. Периоды клеточного цикла в интерфазе. Клеточный цикл эукариот состоит из двух периодов:
Период клеточного роста, называемый «интерфаза», во время которого идет синтез ДНК и белков и осуществляется подготовка к делению клетки.
Период клеточного деления, называемый «фаза М» (от слова mitosis — митоз).
Интерфаза состоит из нескольких периодов: а) G1-фазы (от англ. gap — промежуток), или фазы начального роста, во время которой идет синтез мРНК, белков, других клеточных компонентов; б) S-фазы (от англ. synthesis — синтез), во время которой идет репликация ДНК клеточного ядра, также происходит удвоение центриолей (если они, конечно, есть). в) G2-фазы, во время которой идет подготовка к митозу. [У дифференцировавшихся клеток, которые более не делятся, в клеточном цикле может отсутствовать G1 фаза. Такие клетки находятся в фазе покоя G0.]
Период клеточного деления (фаза М) включает две стадии:
кариокинез (деление клеточного ядра); -> цитокинез (деление цитоплазмы).
В свою очередь, митоз делится на пять стадий. Митоз и его фазы. 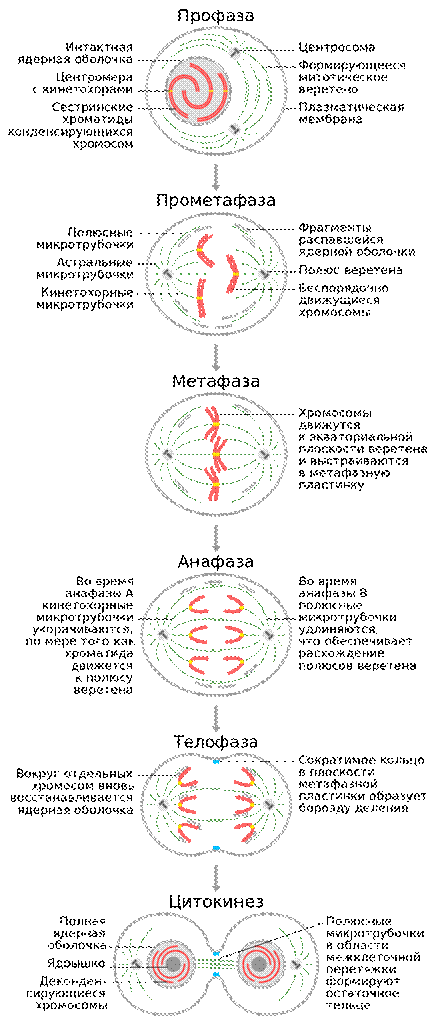
7. Мейоз как механизм полового размножения состоит из 2 последовательных делений с короткой интерфазой между ними.
- Профаза I — профаза первого деления очень сложная и состоит из 5 стадий:
- Лептотена или лептонема — упаковка хромосом, конденсация ДНК с образованием хромосом в виде тонких нитей (хромосомы укорачиваются).
- Зиготена или зигонема — происходит конъюгация — соединение гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами и их дальнейшая компактизация.
- Пахитена или пахинема — (самая длительная стадия) — в некоторых местах гомологичные хромосомы плотно соединяются, образуя хиазмы. В них происходит кроссинговер — обмен участками между гомологичными хромосомами.
- Диплотена или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой. У некоторых животных в ооцитах хромосомы на этой стадии профазы мейоза приобретают характерную форму хромосом типа ламповых щёток.
- Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; центриоли расходятся к полюсам; гомологичные хромосомы остаются соединёнными между собой.
К концу Профазы I центриоли мигрируют к полюсам клетки, формируются нити веретена деления, разрушаются ядерная мембрана и ядрышки
- Метафаза I — бивалентные хромосомы выстраиваются вдоль экватора клетки.
- Анафаза I — микротрубочки сокращаются, биваленты делятся, и хромосомы расходятся к полюсам. Важно отметить, что, из-за конъюгации хромосом в зиготене, к полюсам расходятся целые хромосомы, состоящие из двух хроматид каждая, а не отдельные хроматиды, как в митозе.
- Телофаза I — хромосомы деспирализуются и появляется ядерная оболочка.
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
- Профаза II — происходит конденсация хромосом, клеточный центр делится и продукты его деления расходятся к полюсам ядра, разрушается ядерная оболочка, образуется веретено деления, перпендикулярное первому веретену.
- Метафаза II — унивалентные хромосомы (состоящие из двух хроматид каждая) располагаются на «экваторе» (на равном расстоянии от «полюсов» ядра) в одной плоскости, образуя так называемую метафазную пластинку.
- Анафаза II — униваленты делятся и хроматиды расходятся к полюсам.
- Телофаза II — хромосомы деспирализуются и появляется ядерная оболочка.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки. В тех случаях, когда мейоз сопряжён с гаметогенезом (например, у многоклеточных животных), при развитии яйцеклеток первое и второе деления мейоза резко неравномерны. В результате формируется одна гаплоидная яйцеклетка и три так называемых редукционных тельца (абортивные дериваты первого и второго делений). Значение. У организмов, размножающихся половым путём, предотвращается удвоение числа хромосом в каждом поколении, так как при образовании половых клеток мейозом происходит редукция числа хромосом.
Мейоз создает возможность для возникновения новых комбинаций генов (комбинативная изменчивость), так как происходит образование генетически различных гамет.
Редукция числа хромосом приводит к образованию «чистых гамет», несущих только один аллель соответствующего локуса.
Расположение бивалентов экваториальной пластинки веретена деления в метафазе 1 и хромосом в метафазе 2 определяется случайным образом. Последующее расхождение хромосом в анафазе приводит к образованию новых комбинаций аллелей в гаметах. Независимое расхождение хромосом лежит в основе третьего закона Менделя.
8. Чередование гаплоидной и диплоидной фаз в жизненном цикле представителей разных систематических групп.
Организмам, размножающимся только половым путем, характерно чередование гаплоидной и диплоидной фаз в их развитии. У многих организмов, включая млекопитающих, это чередование имеет регулярный характер, и на нем основано сохранение видовых признаков организмов. Диплоидия способствует накоплению разных аллелей. Напротив, для организмов, которые могут размножаться как половым, так и бесполым путем, характерно чередование (смена) поколений, когда одно или несколько бесполых поколений организмов сменяется поколением организмов, размножающихся половым путем.
Различают первичное и вторичное чередование поколений. Первичное чередование поколений отмечается у организмов, развивших в ходе эволюции половой прогресс, но сохранивших способность к бесполому размножению, и заключается в регулярном чередовании полового и бесполого поколений Оно встречается у животных (простейших), у водорослей и у всех высших растений. У простейших классическим примером первичного чередования поколений является бесполое размножение малярийного плазмодия в организме человека (шизогония) и половое — в организме малярийного комара. У растений половое поколение представлено гаметофитом, бесполое — спорофитом. Механизм первичного чередования заключается в том, что на растениях спорофитного поколения развиваются споры, которые на основе мейоза дают гаплоидные мужские и женские гаметофиты. На последних развиваются спермии и яйцеклетки. Оплодотворение яйцеклетки дает начало диплоидному спорофиту. Таким образом, клетки гаметофита содержат гаплоидный набор хромосом, а спорофита — диплоидный набор, т. е. у растений чередование поколений связано со сменой гаплоидного и диплоидного состояний.
Гаметофит и спорофит на примере жизненного цикла папоротника
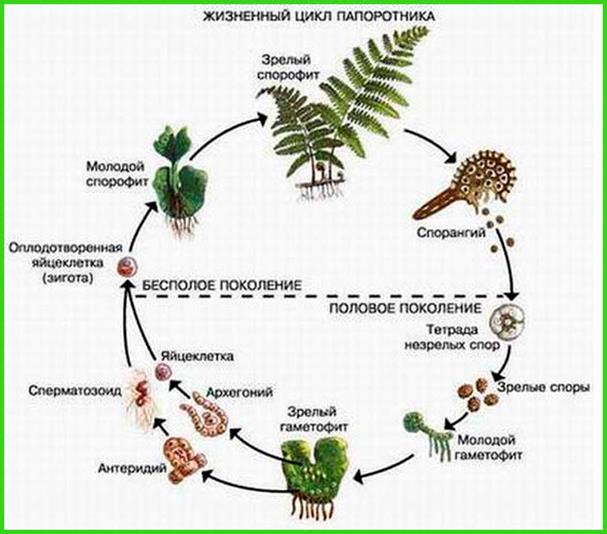
Если проследить за соотношением между спорофитом и гаметофитом у растений разного уровня организации, то можно увидеть, что в ходе эволюции развитию подвергался спорофит, тогда как для гаметофита была характерной редукция. Например, у мхов преобладающим является гаметофит (гаплоидное поколение), на котором живет спорофит. Но уже у папоротникообразных преобладающим является спорофит (диплоидное поколение) в виде хорошо развитого растения со стеблями и корнями, а гаметофит представлен слоем клеток, которые образуют пластину, прикрепляющуюся к почве с помощью ризоидов. Далее, у голосеменных гаметофит уменьшается до небольших количеств клеток, а у покрытосеменных мужской гематофит представлен лишь двумя клетками, женский — семью, тогда как спорофитом у голосеменных являются деревья (сосна, ель и другие), а покрытосеменных — деревья, кустарники, травы.
Между гаметофитом и спорофитом могут быть как сходства по морфологии и продолжительности жизни, так и различия по этим признакам. В первом случае это называют изоморфным чередованием поколений, во втором — гетероморфным.
Вторичное чередование поколений широко встречается у животных. Оно отмечается в формах гетерогонии и метагенеза. Гетерогония заключается в первичном чередовании полового процесса и партеногенеза. Например, у трематод половое размножение регулярно сменяется партеногенезом. У многих других организмов гетерогония зависит от сезона. Так, коловратки, дафнии и тли осенью размножаются путем зигогенеза (путем оплодотворения яйцеклеток и образования зигот), а летом — путем партеногенеза. Метагенез заключается в чередовании полового размножения и вегетативного (бесполового). Например, гидры размножаются обычно почкованием, но при понижении температуры образуют половые клетки. У кишечнополостных на некоторых стадиях развития происходит переход от полового размножения к вегетативному. У некоторых морских кишечнополостных полипоидное поколение правильно чередуется с медузоидным. Для полипоидного поколения характерно размножение так называемой стробиляцией (поперечными перетяжками), для медузоидного — половым путем (оплодотворение яиц, образование личинок и развитие полипов).
9. Развитие половых клеток у животных и человека: сперматогенез.
Сперматогене́з — развитие мужских половых клеток (сперматозоидов), происходящее под регулирующим воздействием гормонов. Одна из форм гаметогенеза. Сперматозоиды развиваются из клеток-предшественников, которые проходят редукционные деления (деления мейоза) и формируют специализированные структуры (акросома, жгутик и пр.). В разных группах животных сперматогенез различается. У позвоночных животных сперматогенез проходит по следующей схеме: в эмбриогенезе первичные половые клетки — гоноциты мигрируют в зачаток гонады, где формируют популяцию клеток, называемых сперматогониями. С началом полового созревания сперматогонии начинают активно размножаться, часть из них дифференцируется в другой клеточный тип — сперматоциты I порядка, которые вступают в мейоз и после первого деления мейоза дают популяцию клеток, называемых сперматоцитами II порядка, проходящих впоследствии второе деление мейоза и образующих сперматиды; путём ряда преобразований последние приобретают форму и структуры сперматозоида в ходе спермиогенеза.
Сперматогенез у человека в норме начинается в пубертатном периоде (около 12 лет) и продолжается до глубокой старости. Продолжительность полного сперматогенеза у мужчин составляет примерно 73—75 дней[1]. Один цикл зародышевого эпителия составляет приблизительно 16 дней. Сперматозоиды образуются в семенниках, а именно в извитых семенных канальцах. Стенка семенного канальца делится базальной мембраной на люминальную и адлюминальную стороны. На люминальной стороне расположены клетки Сертоли (сустентоциты) и предшественники половых клеток (сперматогонии, сперматоциты I и II порядков и сперматиды). Сперматогонии, лежащие непосредственно на базальной мембране извитых семенных канальцев, проходят несколько последовательных стадий митотического деления. Общее количество сперматогоний в яичке мужчины составляет около 1 млрд. Различают две основные категории сперматогоний: А и В. Сперматогонии А, которые делятся митотически, сохраняют способность к делению и поддерживают свою популяцию. Остальные дифференцируются в сперматогоний В, которые «эвакуируются» клеточными контактами сустентоцитов (образуют под основанием половой клетки новый контакт и резорбируют старый). Сперматогония В делится митотически, дифференцируясь в сперматоцит I порядка, вступающий в мейоз. В результате первого деления мейоза образуются две дочерние клетки сперматоциты второго порядка, каждый из которых содержит гаплоидный набор (23 у человека) d-хромосом. Вторичные сперматоциты расположены ближе к просвету канальца. Во втором делении мейоза образуются две сперматиды. Таким образом, в результате деления одной сперматогонии образуются четыре сперматиды, каждая из которых обладает гаплоидным набором хромосом.
В ходе сложного процесса спермиогенеза сперматиды дифференцируются в зрелые сперматозоиды. Дифференцирующиеся сперматиды лежат в углублениях плазматической мембраны клеток Сертоли. При спермиогенезе комплекс Гольджи формирует акросому, содержащую протеолитические ферменты, которые при контакте с яйцеклеткой растворяют участок её блестящей оболочки (zona pellucida).
Сложный процесс сперматогенеза регулируется гонадотропными гормонами гипофиза и стероидными гормонами яичка. После полового созревания гипоталамус начинает выделять гонадотропный рилизинг-гормон, под влиянием которого гипофиз секретирует фолликулостимулирующий гормон (ФСГ), стимулирующий развитие и функционирование клетки Сертоли и лютеинизирующий гормон (ЛГ), стимулирующий клетки Лейдига к выработке тестостерона. Тестостерон оказывает воздействие на развитие клеток Сертоли, а также на предшественники половых клеток (в ассоциации с андроген-связывающим белком, выделяемым клетками Сертоли).
Секреторная активность гипофиза регулируется клетками Сертоли и клетками Лейдига. Тестостерон, выделяемый клетками Лейдига подавляет активность гипофиза к выработке ЛГ и ФСГ. Ингибин и эстрадиол, образующийся в клетках Сертоли, подавляют гипофиз к выработке ФСГ и клетки Лейдига к выработке тестостерона. Морфофункциональное состояние яичка регулируется гормонами аденогипофиза — ФСГ и ЛГ, причём уровень гормонов постоянный, имеются лишь незначительные колебания.
и овогенез. — это процесс формирования яйцеклеток. Его функциями являются обеспечение гаплоидного набора хромосом в ядре яйцеклетки и обеспечение питательных потребностей зиготы. Овогенез в своем проявлении в основном сопоставим со сперматогенезом.
У млекопитающих и человека овогенез начинается еще во внутриутробном периоде (до рождения). Овогонии, представляющие собой мелкие клетки с довольно крупным ядром и локализованные в фолликулах яичников, начинают в фолликулах дифференцироваться в первичные овоциты. Последние формируются уже на третьем месяце внутриутробного развития, после чего они вступают в профазу первого мейотического деления. Ко времени рождения девочки все первичные овоциты уже находятся в профазе первого мейотического деления. Первичные овоциты остаются в профазе вплоть до наступления половой зрелости женского индивида. Когда при наступлении половой зрелости фолликулы яичника созревают, мейотическая профаза в первичных овоцитах возобновляется. Первое мейотическое деление для каждой развивающейся яйцеклетки завершается вскоре перед временем овуляции этой яйцеклетки. В результате первого мейотического деления и неравномерного распределения цитоплазмы одна образовавшаяся клетка становится вторичным овоцитом, другая — полярным (редукционным) тельцем.
Вторичное мейотическое деление у человека наступает, когда вторичный овоцит (развивающаяся яйцеклетка) проходит из яичника в фаллопиеву трубу. Однако это деление не завершается до тех пор, пока во вторичный овоцит не проникает ядерное содержимое сперматозоида, что происходит обычно в фаллопиевой трубе. Когда ядро сперматозоида проникает во вторичный овоцит, то последний делится, в результате чего образуется овотида (зрелая яйцеклетка) с пронуклеусом, содержащим одиночный набор из 23 материнских хромосом. У некоторых других видов образуются яйцеклетки, определяющие как мужской пол, так и женский. Важно подчеркнуть, что здесь происходят также расщепление и рекомбинация генов, основу которой создает расхождение хромосом. Другая клетка, образующаяся в результате второго мейотического деления у человека, является вторым полярным тельцем, не способным к дальнейшему развитию. В это время подвергается делению надвое и полярное (редукционное) тельце. Таким образом, развитие одного овоцита первого порядка сопровождается образованием одной овотиды и трех редукционных телец. В яичниках таким путем на протяжении жизни обычно созревает 300-400 овоцитов, но в месяц созревает лишь один овоцит. В течение дифференциации яйцеклеток формируются мембраны, уменьшается в размере их ядро.
У некоторых видов животных овогенез протекает быстро и непрерывно и приводит к образованию большого количества яйцеклеток.
Вопреки сходству со сперматогенезом овогенез характеризуется некоторыми специфическими особенностями. Питательный материал (желток) первичного овоцита не распределяется поровну между четырьмя клетками, которые образуются в результате мейотических делений. Основное количество желтка сохраняется в одной большой клетке, тогда как полярные тельца содержат очень мало этого вещества. Первые и вторые полярные тельца получают в результате делений те же самые хромосомные наборы, что и вторичные овоциты, но они не становятся половыми клетками. Поэтому яйцеклетки намного богаче питательным материалом по сравнению со сперматозоидами. Особенно сильно это различие проявляется в случае яйцекладущих животных.
Яйцеклетки млекопитающих имеют овальную или несколько вытянутую формуи характеризуются типичными чертами клеточного строения. Они содержат все структуры, характерные для соматических клеток, однако внутриклеточная организация яйцеклетки очень специфична и определяется тем, что яйцеклетка является также и средой, которая обеспечивает развитие зиготы. Одна из характерных особенностей яйцеклеток заключается в сложности строения их оболочек. У очень многих животных различают первичную, вторичную и третичную оболочки яйцеклеток. Первичная оболочка (внутренняя) формируется еще на стадии овоцита. Представляя собой поверхностный слой овоцита, она имеет сложную структуру, т. к. пронизана выростами прилегающих к ней фолликулярных клеток. Вторичная (средняя) оболочка полностью сформирована фолликулярными клетками, а третичная (наружная) образована веществами, представляющими собой продукты секреции желез яйцеводов, через которые проходят яйцеклетки. У птиц, например, третичными оболочками яйцеклеток служат белковая, подскорлуповая и скорлуповая оболочки. Для яйцеклеток млекопитающих характерно наличие двух оболочек. Структура внутриклеточных компонентов яйцеклеток специфична в видовом отношении, а иногда имеет даже индивидуальные особенности.
У мужчин образование и выделение гамет - процесс непрерывный. Он начинается с наступлением половой зрелости и продолжается в течение всей жизни. У женщин же, гаметогенез - это периодический процесс, повторяющиеся примерно через каждые ~28 дней. Во время каждого периода происходят структурно-функциональные изменения всей системы репродукции. Процесс этот называется овариально-менструальным циклом. В овариально-менструальном цикле участвуют яичники (овариальный период) и матка (маточный период). Все процессы овариально-менструального цикла управляются посредством гормонов яичников, секреция которых в свою очередь управляется гипофизарными гонадотропинами.
| Сперматогенез | Овогенез |
| Фаза размножения происходит только с момента полового созревания и продолжается в течение всей жизни | Фаза размножения происходит только в эмбриональном периоде и непродолжительное время после рождения |
| Фаза роста сразу следует за размножением, короткая | Фаза роста очень длительная, делится на малый рост и большой рост |
| Фаза созревания характеризуется равномерным делением сперматоцитов | Фаза созревания характеризуется неравномерным делением овоцитов: образуется 1 яйцеклетка и 3 редукционных тельца |
| Есть фаза формирования | Фаза формирования отсутствует |
| «Экономичность» сперматогенеза: из одной сперматогонии образуется 4 сперматозоида | «Расточительность» овогенеза: из одной овогонии образуется 1 крупная яйцеклетка и 3 мелких редукционных тельца |
| Продолжается в течение всей жизни мужчины | Прекращается после менопаузы |
10. Основные черты эмбриогенеза анамний. Черты сходства и различия в эмбриогенезе анамний.
Анамнии (лат. Anamnia) или низшие позвоночные — парафилетическая группа, включающая позвоночных животных, не имеющих зародышевых оболочек. В отличие от амниотов, у анамний в процессе эмбрионального развития не возникает зародышевой оболочки — амниона и особого зародышевого органа — аллантоиса. Анамнии связаны в своём существовании с водной средой, в которой они проводят либо всю жизнь, либо начальные стадии (яйцевые и личиночные). К анамниям относятся все позвоночные, за исключением амниот, то есть бесчелюстные и различные группы рыб, а также ам
|
|
|
