 |


Эволюция растительности Земли
|
|
|
|
ЛЕКЦИЯ № 22
ТЕМА: ЭВОЛЮЦИЯ РАСТЕНИЙ.
Эволюция царства растений началась с архейской эры около 3500 млн. лет назад). В этот период на Земле отмечено появление синезеленых водорослей. Эти водоросли принадлежат к группе цианобактерии, поскольку в их клетках отсутствует оформленные ядра. Таким образом, их можно отнести к прокариотам (доядерным организмам). Среди синезеленых водорослей были одно- и многоклеточные организмы, имеющие возможность осуществлять фотосинтез. Благодаря процессу фотосинтеза, в атмосферу нашей планеты начал поступать кислород, необходимый для жизнедеятельности аэробов.

Позднее в протерозойской эре (около 2600 млн. лет назад) Землей завладели красные и зеленые водоросли. Их господство распространилось и на палеозойскую эру (примерно 570 млн. лет назад). Только в поздний палеозой (силурийский период) отмечена жизнедеятельность на планете древнейших высших растений – риниофитов, или псилофитов. У этих растений были побеги, но отсутствовали корни и листья. Размножение риниофитов происходило спорами. Они обитали на суше или частично в воде.
В существовании нашей планеты новая эра началась с появления высших, или наземных растений. Около 400- 360 млн. лет назад в девонском периоде палеозойской эры на фоне преобладания на Земле риниофитов и водорослей появились первые папоротникообразные (папоротники, хвощи, плауны) и моховидные растения. Они относятся к высшим споровым растениям. Благодаря распространению растений на суше, появились и новые наземные виды животных. Сочетанное изменение в ходе эволюции форм растений и животных обусловило огромнейшее разнообразие жизни на Земле. Облик планеты изменился коренным образом. Прикрепленный образ жизни растения на суше привел к появлению расчленения растения на корень, стебель и лист, а также к возникновению опорных тканей и сосудистой проводящей системы. Самые первые наземные растения были маленьких размеров. Они поглощали воду посредством ризоидов, как сохранившиеся по сей день на Земле - мхи.
|
|
|
В цикле их развития преобладало гаплоидное поколение (гаметофит). Постепенно появлялись более крупные формы растений - папоротникообразные, у которых образовались сложные специализированные органы – корни с корневыми волосками. В цикле развития этих растений на первый план выступает диплоидная фаза - спорофит, являющийся непосредственно самим растением, тогда как гаметофит – это заросток, который выглядит как клубенек у хвощей и плаунов и как небольшая пластинка в форме сердца у папоротников. Так осуществлялся постепенный переход от гаплоидного поколения к более совершенному - диплоидному. В палеозойскую эру папоротникообразные были громадными растениями, господствовавшими на суше. Однако для их размножения была необходима вода, что ограничивало территорию их существования местностями с повышенной влажностью.
В каменноугольном периоде, который длился с 360 до 280 млн. лет назад, доказано появление на нашей планете семенных папоротников, которые в дальнейшем стали родоночальниками всех голосеменных растений. В это время полностью исчезают риниофиты из-за невозможности конкурировать с более развитыми растениями. А господствовашие тогда огромнейшие древовидные папоротникообразные после отмирания образовали залежи каменного угля.
В следующем пермском периоде палеозойской эры на Земле появились древние голосеменные растения. Древовидные папоротникообразные постепенно вымирают, а им на смену приходят семенные и травянистые папоротники, завладевая сушей. Особенностью голосеменных растений является то, что размножение их осуществляется семенами, не имеющими защиты в виде стенок плода, так как цветков и плодов эти растения не образуют. Половое размножение этих растений осуществлялось независимо от капельно-водной среды. А их появление в ходе эволюционных метаморфоз было обусловлено перепадами влажности и температуры и изменением рельефа Земли вследствие поднятия суши, то есть появления горных массивов.
|
|
|
Мезозойская эра наступила около 240 млн. лет назад. В триасовом периоде мезозоя появились современные голосеменные, а в юрском периоде зародились первые покрытосеменные растения. Но господствующие позиции на планете сохранились за голосеменными растениями. Это эра вымирания древних папоротникообразных, не выдерживающих естественного отбора. В процессе появления покрытосеменных растений произошла череда ароморфозов. Во-первых, сформировался цветок – трансформированный побег, приспособленный для образования спор и гамет. Опыление, оплодотворение и образование зародыша и плода происходило непосредственно в цветке. Во-вторых, для лучшей защиты и распространения семена покрытосеменных растений были окружены околоплодником. Для этих растений характерно половое размножение. К покрытосеменным относятся травянистые растения, деревья и кустарники. Разнообразные видоизменения вегетативных органов (корня, стебля, листа) отмечаются у разных видов растений. Эволюционные изменения покрытосеменных растений происходили за относительно короткий срок, поэтому для них свойственна высокая эволюционная пластичность. Огромное значение в протекании эволюционных преобразований сыграли насекомые-опылители. Покрытосеменные растения более продуктивно осваивают окружающую среду и завоевывают новые территории, благодаря своим особенностям строения и способности формировать сложные многоярусные сообщества.
В кайнозойской эре, наступившей приблизительно 70 млн. лет назад, на нашей планете стали господствовать существующие и в настоящее время покрытосеменные и голосеменные растения, тогда как высшие споровые растения регрессировали.

Как произошел корень
Водоросли, как живущие в морях солоноводные, так и живущие в пресных водах озер, рек и пр., впитывают воду всей поверхностью. Каждая их клетка, граничащая с внешней средой, представляет собою осмотический аппарат, всасывающий растворы минеральных солей и газов и отдающий растворы органических соединений, напр., органических кислот и их солей. Даже те водоросли, которые живут на коре деревьев или на скалах, одевая их зелеными (плеурококки, празиола) или красными (трентеполия) пятнами, поглощают воды росы или дождя всей поверхностью. Если же росы и дождя нет, то они на время замирают, чтобы снова стать деятельными, лишь только дождь коснется их снова. Ясно, что у них и не может быть специальных органов всасывания воды.
|
|
|

Рис.3 Строение морской водоросли – ламинарии
Мхи начинают свою жизнь стадией предростка, построенного наподобие водоросли и погруженного в воду. В редких случаях, когда предросток мха развивается на обсыхающей почве, отдельные его ветви оказываются в земле, и на них развиваются длинные трубчатые клетки, называемые ризоидами, которые всасывают воду, в то время как верхняя, находящаяся на свету часть предростка ее испаряет. В виде исключения имеется несколько водорослей, имеющих как бы два конца: всасывающий и испаряющий; иначе говоря, им свойственны явления полярности. Нижний полюс погружен в почву и поглощает воду, верхний обращен к солнцу и ее теряет.Такова, например, маленькая водоросль ботридий (Botrydium granulatum), растущая на отмелях и на сырых пашнях. Вся она имеет грушевидную форму, а нижняя ее часть вытянута в узкую ветвистую у основания трубочку. Внутренних перегородок у ботридия нет, строение его сифонное, и вся протоплазма, если освещение слишком сильное, перекочевывает свободно вниз и так же свободно подымается наверх, если небо пасмурно.

Рис.4. Полярное строение ботридия
Воду ботридий поглощает нижней своей частью, разветвления которой называются ризоидами. Полярность, т. е. противоположение испаряющей части растения и части, всасывающей воду,— вот первый импульс к образованию первых корнеподобных органов. Гаметофиты мхов, даже наиболее крупных (исключение — сфагновые мхи и мхи подводные), поглощают воду также ризоидами, т. е. отдельными клетками, покрывающими нижнюю часть стебля. Никакого подобия настоящего корня у мхов нет, как вообще его нет ни у одного гаметофита гаплоида. Зародыш мха, развивающийся из оплодотворенной яйцеклетки и поэтому диплоидный, имеет палочкообразную или овальную форму, он или вовсе не дифференцирован, или же имеет слабо отличающуюся пяту, с помощью которой присасывается к тканям стебля растения матери и из них всасывает водные растворы. У древнейшей группы наземных растений, у псилофитов, корней не было вовсе, воду они вбирали, так же как и мхи, с помощью ризоидов. Только у рода астероксилон, побеги которого слабо напоминают плауновые растения, есть вильчато разветвленные, отходящие от корневища отростки, напоминающие корни. У плаунов, хвощей и у всех папоротников, как у растений диплоидных, корни уже вполне развиты. Они всегда невелики, отходят от корневища или от коры стебля и неглубоко погружаются в почву. Ввиду того, что все эти растения жили и живут во влажных местностях, они могут довольствоваться поверхностными корнями.
|
|
|



Рис. 5. Строение папоротникообразных (акцент на наличие корней)
Чем крупнее растение, чем более развита его испаряющая крона, тем более повышается потребность в глубоко зарывающейся в почву мощной корневой системе.
У голосеменных растений впервые появляется стержневой корень, закладывающийся уже в период развития зародыша в семени. Такой корень дает возможность молодому растению быстро углубиться в почву и распределить боковые мочки в таком слое почвы, который наилучше обеспечивает водоснабжение растения. Однако у корня есть и еще функция — это функция прикрепления к почве, функция механическая. Уже у крупных морских водорослей типа ламинарии от нижней части стебля отходят мощные ризоиды, впивающиеся в камни и удерживающие водоросль на определенной глубине, как бы ни били ее волны. Здесь функция всасывания не имеет значения, все дело в механическом сопротивлении среде.
Корни наших деревьев должны отвечать огромному сопротивлению, так как ветер очень сильно давит на обширную площадь кроны; дерево гнется, иногда ломается, а корни не поддаются, сохраняя связь менаду деревом и почвою. Зато при условиях, неблагоприятных развитию корневой системы, ветровал неизбежен. У растений, живущих на очень сухих почвах, поражает длина корней, часто во много раз превышающая в вышину наземные части растения. Напомним, что анатомически корень состоит из основного цилиндра, коры, конуса нарастания и защищающего этот конус чехлика. Кора на некотором расстоянии от конца корня несет зону корневых волосков, усиливающих всасывание. Осевой цилиндр содержит в себе проводящие воду сосуды, через которые она поступает в стебель. Клетки коры развивают колоссальное осмотическое давление, обеспечивающее движение воды в сосудах. Если корень находится в симбиозе с грибками, образуя микоризу, то корневые волоски, как правило, отсутствуют.
|
|
|
Корень постоянно растет. Подвергаясь действию внешних факторов, как то: силе тяжести, свету, теплу, влажности, присутствию кислорода воздуха и химизму почвенного раствора,— корень растет неравномерно, искривляется и проникает в наиболее благоприятные для него слои почвы. Таков результат длительной, постоянно поддерживающейся борьбы за существование.
Развитие стебля
Основная форма стебля — форма колонны — так проста, что в этом отношении никакой особой эволюции от крупной морской водоросли лессонии через стволы плауновых деревьев каменноугольной эпохи к стволам крупных хвойных и лиственных деревьев не замечается. Наоборот, анатомия стебля дает очень сложную картину постепенного усложнения и усовершенствования проводящего аппарата. У бурых водорослей в центральной части их стебля мы находим наличие длинных трубчатых клеток, имеющих анастомозы и сообщающихся одна с другой помощью ситовидных пластинок. Последнее — не что иное, как поперечные перегородки, отделяющие одну клетку от другой. Протоплазмы же их проходят через поры сит, неся с собой питательные вещества. Ствол массивен, ассимиляция происходит в поверхностных слоях ткани, внутренние части, затененные наружными, будут голодать, если их не будет пронизывать система трубочек с движущимися растворами внутри.
Даже у грибов в их более массивных стеблевидных образованиях, каковы, например, шнуровидные ризоморфы опенка, разрастающиеся под корою старых пней, внутренние клетки принимают на себя роль проводящей ткани и функционируют соответственно, представляя собою пучок длинных нитей, окруженных мелкоклеточной плотной корой.
У мхов, в их тонких стеблях, мы всегда находим в центре проводящий цилиндр из узких тонких трубчатых клеток, образующих точно пригнанные вертикальные ряды. Поперечные разрезы шлифов стеблей псилофитов показывают сразу, что эти первые растения суши были построены сложнее мхов, хотя и сходны с ними. У ринии в центре стебля находился участок толстостенных клеток, проводивших воду от корневища к верхушкам побегов. Участок этот был окружен кольцом многочисленных тонкостенных трубчатых клеток, проводивших питательные вещества от зеленых верхних частей растения в корневище. У астероксилона на продольных шлифах видны водоносные клетки — трахеиды с характерными кольчатыми или сетчатыми утолщениями на стенках. Совокупность всех таких проводящих клеток, имеющая вид внутреннего шнура, пролегающего между более рыхлой мякотью коры стебля, получила наименование стеле, или столба.
Эволюция стеле папоротников, приведшая к большому разнообразию и строения, и расположения, в тех случаях, когда их много, привела в конце концов к выработке наиболее рационального построения древесины и коры у цветковых растений. Если у папоротников древесина всегда состоит из водоносных клеток или трахеид, то у хвойных в их молодых тканях уже есть небольшие спиральные сосуды (в протоксилеме), у гнетовых их значительно больше и они принимают некоторое участие также и в строении вторичной древесины.
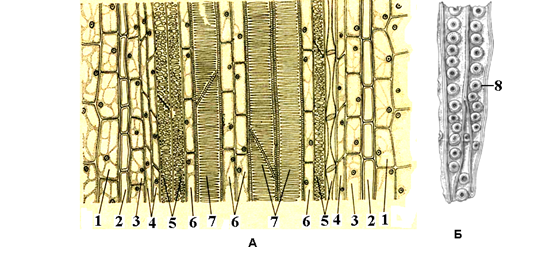
Рис.6. Проводящий пучок папоротника орляка на продольном срезе (А), Б – трахеиды сосны: 1 – паренхима стебля, 2 – эндодерма, 3 – перицикл, 4 – клетки-спутницы, 5 – ситовидные трубки, 5 – древесинная паренхима, 7 – лестничные сосуды, 8 – окаймленные поры.
У однодольных не развит еще камбий, т. е. специальная зародышевая или образовательная ткань проводящих пучков, что мешает им срастаться вместе и расти в толщину. Поэтому у них строение стволов менее плановое, менее совершенное, чем у двудольных растений, дающих благодаря камбию полное разделение ствола па кору и древесину. Таким образом, процесс эволюции охватывает все детали строения стебля, делая его достаточно совершенным орудием как механической крепости растения, так и физиологической связи между двумя активными системами органов растительной жизни, именно между листовой и корневой системами
Стебель представляет собой ось побега. Он имеет метамерное строение и состоит из узлов и междоузлий. Нарастает стебель не только за счет верхушечного, но нередко и за счет вставочного роста. Этим стебель существенно отличается от корня. Стебли чрезвычайно разнообразны по форме, они могут быть четырехгранными, многогранными, плоскими в сечении, однако наиболее распространенная форма - цилиндрическая.
Основные функции стебля опорная и проводящая. Стебель морфологически и функционально связывает между собой основные органы питания -корень и листья -это орган-посредник. При размножении растений черенками мы практически используем важное свойство стебля регенерировать недостающие органы. К этим основным функциям присоединяются многие другие, а иногда они гипертрофированны и выступают на первый план: это запасающая функция, защитная функция (когда стебель несет колючки), молодые стебли, имеющие под эпидермой хлоренхиму, активно участвуют в фотосинтезе. Стебли растений резко различаются по продолжительности жизни. Прежде всего выделяют однолетние и многолетние растения, травянистые и древесные жизненные формы. Многолетние стебли древесных растений называют стволами, у кустарников отдельные стебли именуются стволиками.
Для стебля характерна сложная система меристем: верхушечных, боковых и вставочных.
В результате деятельности первичных меристем складывается первичное строение стебля. У некоторых растений, например, у большинства однодольных, оно сохраняется на протяжении всей жизни. В том случае, если закладывается прослойка камбия, начинается вторичное утолщение стебля. Вторичное утолщение стебля свойственно лишь немногим однодольным - преимущественно древовидным представителям семейства лилейных: драценам, юккам, алоэ.
Первичное строение стебля. В стебле первичного строения, как и в корне, различают центральный осевой цилиндр (стелу) и первичную кору (ПК). Но соотношение этих тканей совершенно иное: большую часть поперечного сечения занимает центральный цилиндр, имеющий нередко в центре крупную воздушную полость (ВП). Кроме того, в отличие от корня, первичная кора стебля покрыта типичной эпидермой с устьицами (в корне экзодерма). Нередко непосредственно под эпидермой залегают механические ткани (колленхима или склеренхима), вынесенные у стебля на периферию. Эти ткани встречаются в различных сочетаниях с хлорофиллоносной, выделительной и неспециализированной паренхимой. Граница между корой и стелой в стебле выражена значительно менее четко, чем в корне.
Наиболее сложное строение имеет стела. Здесь же наблюдается и максимальное разнообразие различных вариантов строения. По характеру расположения проводящих тканей различают 1) стебли пучкового типа (у которых эти ткани закладываются в виде отдельных пучков) и 2) стебли непучкового типа: в этом случае проводящие ткани располагаются сплошным кольцом.
Флоэма в пучках закладывается снаружи (экзархно) и развивается центростремительно.
Ксилема же закладывается эндархно и развивается центробежно.
Таким образом, в стебле эти ткани развиваются друг навстречу другу.
Вторичное утолщение стебля. Если прокамбий всецело расходуется на образование первичных проводящих тканей вторичное утолщение не происходит, если же остается меристематически деятельная прослойка, то она со временем превращается в камбий и начинается вторичное утолщение. При вторичном утолщении клетки камбия активно работают, откладывая наружу слои вторичной флоэмы, а внутрь? слои вторичной ксилемы. То есть в развитии тканей происходит полная инверсия (поворот на 180град).
Одновременно с началом проявления камбиальной активности происходят изменения и в покровных тканях. Эпидерма не выдерживает давления нарастающих тканей, распирающих ее изнутри, она разрывается и слущивается. Начинает свою работу феллоген (пробковый камбий). На смену эпидерме приходит вторичная покровная ткань - перидерма.
Развитие стелы
Стела – совокупность первичных проводящих пучков вместе с заключенными между ними прочими тканями и примыкающим к коре перициклом.

Рис.7. Эволюция типов стелы (по Бавтуто, Еремин, 1996): 1 – протостела; 2 – актиностела; 3 – плектостела; 4 – эктофлойная сифоностела; 5 – артростела; 6 – амфифлойная сифо- ностела; 7 – диктиостела; 8- эустела; 9 – атактостела.
В 1886 году Ф. Тигем и А.Дулио впервые ввели понятие «стела», заложив основы стелярной теории – учения о типах строения и закономерностях эволюции центрального цилиндра всех высших растений.
Наиболее древний и примитивный тип стелы – протостела – присущ первым наземным растениям (риниофитам). Последующая эволюция стелы была связана с увеличением поверхности проводящих тканей. Из протостелы развилась актиностела, ксилема которой на поперечном срезе имеет вид звезды. Данный тип стелы также характерен для примитивных растений (плауновидных, вымерших хвощевидных). Переход от протостелы к актиностеле связан с возникновением пучков, идущих в боковые органы; кроме того, в актиностеле проводящие ткани имеют большую поверхность соприкосновения с окружающими живыми тканями, что способствует лучшему проведению веществ.
Результат дальнейшей специализации актиностелы – появление плектостелы, характерной для осевых органов плауновидных. В плектостеле ксилема представлена отдельными участками, расположенными параллельно или радиально друг к другу. В процессе эволюции протостела дала также начало сифоностеле. Сифоностела имеет трубчатое строение и обладает сердцевиной. Возникновение сифоностелы обеспечило существование более крупных растений, т. к. расположение ксилемы на периферии сделало стебли более прочными. Возникновение сердцевины увеличило поверхность соприкосновения проводящих тканей с живыми клетками, а также создало возможность запасания питательных веществ. Известны два типа сифоностелы:
· эктофлойная (флоэма охватывает ксилему снаружи),
· амфифлойная (флоэма окружает ксилему снаружи и изнутри).
Дальнейшее эволюционное усложнение стелы связано с развитием крупных боковых органов. В местах их отхождения от стебля в стеле, в области узлов, образуются большие листовые лакуны (прорывы), заполненные паренхимой. Вследствие этого сифоностела паренхимными полосами (радиальными лучами), идущими от сердцевины к коре в радиальном направлении, рассекается на отдельные пучки, сохраняющие общее положение по цилиндрической поверхности (на поперечном срезе – кольцом).
Возникают диктиостела, артростела и эустела.
Диктиостела (характерна для папоротников, например, для орляка, щитовника, кочедыжника) имеет вид сетчатой трубки. Она возникает из амфифлойной сифоностелы вследствие образования большого количества листовых прорывов.
Эустела встречается у семенных растений и составлена из коллатеральных открытых пучков, расположенных по кругу.
Артростела (характерна для хвощей) является разновидностью эустелы, представлена закрытыми пучками, расположенными вокруг центральной полости стелы и соединяющимися в узлах.
Последнее звено в эволюции стелы стебля представляет собой атактостела однодольных растений. Ей присущи закрытые коллатеральные пучки, сложная картина прохождения пучков по стеблю (дугообразные индивидуальные листовые следы) с их разбросанным расположением по всему поперечному сечению стебля. Эволюция типов стелы показана.
Типы ветвления стебля
· дихотомическое (плаун, хвощи, папоротники, мохообразные)
· моноподиальное (ель, сосна, кипарис)
· симподиальное (черемуха, яблоня, липа, осина)
· ложнодихатомическое (клен, сирень, чубушник): когда две пазушные почки при супротивном листорасположении развиваются одновременно
· кущение: крупные боковые побеги вырастают из самых нижних почек над или под землей
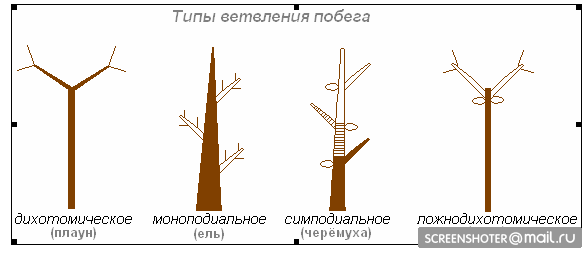
При моноподиальном ветвлении боковые побеги отстают в росте от главного, меристема которого остается работоспособной в течение многих сезонов. При симподиальном ветвлении меристема главного побега образует цветок или отмирает, а рост растения в высоту осуществляют боковые почки, из которых развиваются новые побеги. Наиболее часто встречается такой тип симподиального ветвления, как монохазий, при котором нарастание осуществляется за счет одного бокового побега, который через какое-то время сменяется следующим боковым побегом, и т. д. При развитии дихазия (ложнодихотомическое ветвление) активируется не одна, а две боковых почки, которые продолжают рост побега. Такой тип ветвления характерен для сирени, омелы и многих гвоздичных.
Происхождение листа
У морских и пресноводных водорослей окрашено более или менее равномерно все растение. Изредка лишены хлорофилла ризоиды, да при большой толщине самой водоросли внутренние ткани могут быть очень бедны хромопластами. У мхов дело обстоит уже иначе, гаметофит мха может иметь зеленый стебель, часто плоский или крылатый, или же крыло стебля превращено в листья, морфологически вполне обособленные, а у листостебельных мхов даже и с водопроводящей жилкой. Спорофит мха содержит мало хлорофилльных зерен, которые находятся у него частью в клетках тканей коробочки, частью в подпирающем коробочку апофизе. Настоящих листьев у него нет вовсе. У мелколиственных папоротникообразных (Lycopsida), т. е. у хвощей и плаунов, листья принадлежат уже спорофиту и имеют нормальное строение, но они очень малы и работа их невелика. У хвощей ассимиляция углерода производится главным образом стеблями и веточками, листья в ней почти не участвуют, их значение иное, они охраняют от света растущую зону каждого междоузлия, чем задерживают в ней дифференцировку тканей, а отсутствие дифференцировки сохраняет неизменною зародышевую ткань, главная функция которой — деление клеток и рост всего органа. Первые листья, которые вполне оправдывают такое их обозначение, — это листья некоторых псилофитов (род Asteroxylon), обладавшие, по-видимому, уже вполне развитым устьичным аппаратом. Листья ископаемых и современных плаунов следуют за ними. Эти листья все еще примитивные: у них нет завершенного разделения на черешок и пластинку, нет развитой сети жилок; анатомически оба основных аппарата листа, фотосинтетический — палисадная паренхима — и транспирационный — паренхима губчатая, выражены несовершенно, как несовершенно и различие между верхней и нижней сторонами листа.

Рис. 9. Листья плауна
Правда, подобные листья встречаются и у растений, принадлежащих к высоко организованным семействам, но там это результат опрощения, вызываемого внешними условиями, здесь же — первичное строение. У папоротникообразных, названных Джеффреем Pteropsida, листья крупные, с хорошей дифференцировкой на черешок и пластинку. Их примитивность сказывается главным образом в том, что они растут верхушкою, а не основанием, как листья цветковых. Такой способ роста позволяет им развивать нередко добавочный рост отдельных частей листа, курчавость краев и другие уродливости.
Воду ботридий поглощает нижней своей частью, разветвления которой называются ризоидами. Полярность, т. е. противоположение испаряющей части растения и части, всасывающей воду,— вот первый импульс к образованию первых корнеподобных органов. Гаметофиты мхов, даже наиболее крупных (исключение — сфагновые мхи и мхи подводные), поглощают воду также ризоидами, т. е. отдельными клетками, покрывающими нижнюю часть стебля. Никакого подобия настоящего корня у мхов нет, как вообще его нет ни у одного гаметофита гаплоида. Зародыш мха, развивающийся из оплодотворенной яйцеклетки и поэтому диплоидный, имеет палочкообразную или овальную форму, он или вовсе не дифференцирован, или же имеет слабо отличающуюся пяту, с помощью которой присасывается к тканям стебля растения матери и из них всасывает водные растворы. У древнейшей группы наземных растений, у псилофитов, корней не было вовсе, воду они вбирали, так же как и мхи, с помощью ризоидов. Только у рода астероксилон, побеги которого слабо напоминают плауновые растения, есть вильчато разветвленные, отходящие от корневища отростки, напоминающие корни. У плаунов, хвощей и у всех папоротников, как у растений диплоидных, корни уже вполне развиты. Они всегда невелики, отходят от корневища или от коры стебля и неглубоко погружаются в почву. Ввиду того, что все эти растения жили и живут во влажных местностях, они могут довольствоваться поверхностными корнями.

Рис. 9.Лист папоротника
У цветковых мы имеем колоссальное разнообразие листовых форм и строений. Сеть жилок в своей более простой форме дает очень мало анастомозов. Главные жилки располагаются параллельно одна другой, и при перерезке любой из них примыкающий к верхней части перерезанной жилки участок ткани засыхает. Более прогрессивный тип жилкования — сетчатый. Если перерезать одну из главных жилок у листа этого типа, то ткань кругом остается жива, так как получит воду обходным путем, через боковую сеть. Наиболее развитыми являются листья растений из семейства бобовых, появившегося сравнительно поздно. Лист бобовых состоит из листового основания, отходящих от него прилистников, общего черешка и перисто или дланевидно расположенных пластинок — листочков. Благодаря находящимся на черешочках сочленениям листовые пластинки бобовых могут перемещаться под влиянием большей или меньшей напряженности солнечных лучей и становиться под определенным углом к плоскости падения последних, регулируя таким образом интенсивность освещения. Для ассимиляции особенно благоприятны лучи определенной напряженности, при более сильном освещении хлорофилл разрушается быстрее, чем восстанавливается, при более слабом не хватает энергии. Такие сложные листья, как листья гороха, акации и массы других бобовых, с свободным движением отдельных частей, с организованным отводом продуктов ассимиляции из тканей (мякоти) в ситовидные трубки и пр., являются наиболее совершенным выражением эволюции листа.

Рис.11. Листья мимозы
От листа плауновых до листа гороха пройден сложный и долгий путь, приведший к выработке прекрасного пластичного, сообразно условиям среды, аппарата фотосинтеза; фотосинтез же, как известно, — это главный физиологический процесс зеленых растений.
|
|
|
