 |


Биосинтез нуклеиновых кислот Репликация
|
|
|
|
Биосинтез нуклеиновых кислот Репликация
Полимеризация дочерней ДНК на матрице ДНК приводит к ее удвоению или
репликации. Синтез ДНК протекает в ядре в S-фазу клеточного цикла и предшествует делению клетки.
Для реализации репликации необходимы: матрица – расплетенная цепь ДНК, субстраты, участвующие в полимеризации ДНК (нуклеозидтрифосфаты), ферменты, катализирующие этот процесс, ионы Mg2+, а также белковые факторы, обеспечивающие деспирализацию двухнитевой ДНК.
Механизмы репликации одинаковы для прокариот и эукариот.
Синтез ДНК происходит одновременно на обеих цепях ДНК. В основе – принцип комплементарности. Процесс является полуконсервативным, так как по завершении репликации каждая дочерняя молекула ДНК содержит одну родительскую и одну вновь синтезированную цепь.
Репликон – единица генома, в которой содержатся точка начала репликации (origin) – точка ori – последовательность ДНК, в которой инициируется процесс удвоения ДНК, и точка окончания репликации (terminus) – сегмент ДНК, в котором процесс удвоения ДНК останавливается. Репликативная вилка – область репликации ДНК, перемещающаяся от ori вдоль родительской ДНК, которая расплетается и служит матрицей для синтеза дочерней ДНК. У прокариот ДНК имеет форму кольца и содержит один репликон. В ori-сайте (точка начала репликации) цепи расходятся и образуется две репликативных вилки, движущихся в противоположных направлениях. У эукариот имеется большое число репликонов и ori-сайтов,
и репликация проходит одновременно на многих участках ДНК.
Процесс синтеза ДНК включает стадии: инициации, элонгации и терминации.
Этапы репликации у прокариот.
Синтез ДНК у прокариот осуществляют ДНК-полимеразы: ДНК-полимераза I, которая участвует в репликации и репарации; ДНК-полимераза II – вероятно, участвует в репарации;
|
|
|
ДНК-полимераза III – основной фермент репликации.
Инициация репликации. Репликация начинается с расплетения двойной спирали ДНК и образования репликативной вилки. Это осуществляется при помощи ферментов хеликаз, которые перемещаются вдоль цепей ДНК и раскручивают их. Процесс расплетения двойной спирали ДНК является энергозависимым и требует затраты АТФ. Далее SSB-белки (single strand binding) специфично связываются с одноцепочечной ДНК и препятствуют образованию двойных спиралей (ренатурации ДНК) и шпилечных структур.
Интенсивное раскручивание ДНК может приводить к образованию дополнительных супервитков, которые «снимают» топоизомераты (гираза).
Праймаза комплементарно из рибонуклеозидтрифосфатов синтезирует на матрице ДНК праймер, или РНК-затравку – короткий фрагмент РНК (10-12 нуклеотидов), 3’-ОН-конец которого используется для дальнейшего синтеза ДНК ключевым ферментом репликации ДНК у прокариот - ДНК-полимеразой III.
Элонгация репликации. Субстратами для синтеза дочерних цепей являются четыре дезоксинуклеозидтрифосфата (дНТФ): дАТФ, дТТФ, дЦТФ, дГТФ. С 3’-ОН-конца праймера ДНК-полимераза III начинает синтезировать новую цепь ДНК присоединением каждого последующего дНТФ только к 3’-ОН-концу имеющейся цепи с выделением пирофосфата:
НТФ + (НМФ)n → (НМФ)n+1 + ФФН
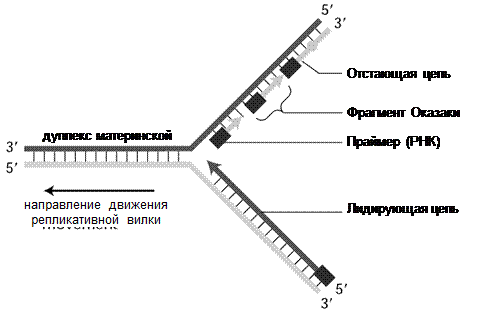
Синтез дочерних цепей ДНК идет в направлении 5’→ 3’ одновременно на обеих цепях матрицы ДНК.
Однако, так как цепи ДНК антипараллельны, а ДНК-полимеразы синтезируют ДНК
присоединением каждого последующего нуклеотида только к 3’-концу имеющейся цепи, в 1960-х гг. Рейжи Оказаки экспериментально доказал, что синтез одной цепи ДНК происходит фрагментарно, прерывисто. Эти фрагменты были названы фрагментами Оказаки.
|
|
|
Т. о. синтез одной цепи происходит непрерывно от 5’- к 3’-концу в направлении движения репликативной вилки и необходим только 1 акт инициации и один праймер. Эта цепь называется лидирующей (рис. 33).
Синтез второй цепи ДНК, которая называется Отстающей, также происходит от 5’- к 3’- концу, но в противоположном направлении репликативной вилки, инициируется многократно, т. о. синтезируется много праймеров и фрагментов Оказаки. У прокариот фрагменты Оказаки имеют длину около 1000 нуклеотидов, у эукариот – 150-300 нуклеотидов.
ДНК-полимераза III синтезирует цепь ДНК до следующей затравки (рис. 34). Далее ДНК-полимераза I удаляет праймер с 5’-конца и одновременно замещая рибонуклеотиды дезоксирибонуклеотидами, а ДНК-лигаза сшивает фрагменты Оказаки в непрерывную цепь.
При включении неправильного нуклеотида ДНК-полимеразы I и III удаляют ошибочный нуклеотид с 3’-конца, гидролизуя фосфодиэфирную связь.
 В результате образуются дочерние цепи, комплементарные и антипараллельные цепям материнской ДНК.
В результате образуются дочерние цепи, комплементарные и антипараллельные цепям материнской ДНК.
|
|
|
