 |


Репарация неспаренных оснований.
|
|
|
|
• Во время репликации ДНК происходят ошибки спаривания.
Неправильное спаривание может затронуть только дочернюю нить ДНК; матричная нить в процессе репликации остается неизменной.
• Следовательно, система репарации должна проводить замену некомплементарных оснований только на дочерней цепи.
• Клетки используют различие в структуре матричной и дочерней нитей.
• Материнская нить ДНК несет метилированные аденины, а в дочерней нити до окончания репликации аденины еще не метилированы.
• Пока они остаются неметилированными, клетки исправляют ошибки.
• 
17/ Чтобы залечить имеющую повреждение ДНК во время репликации, клетка прибегает к следующему приему.
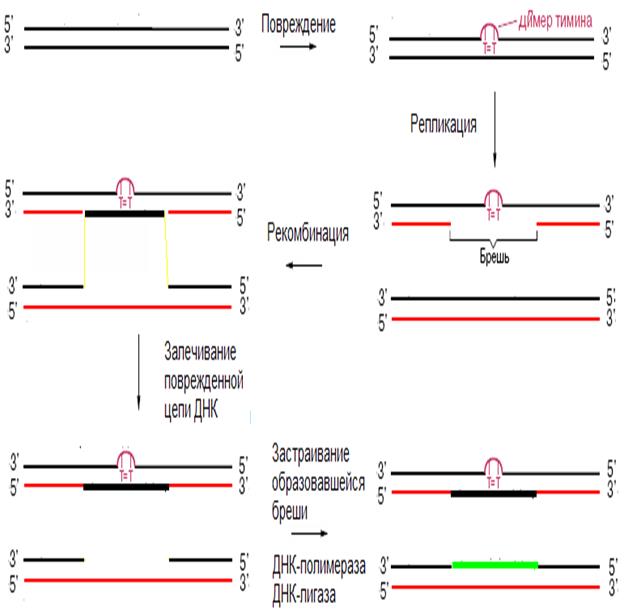
SOS-репарация
• SOS-репарация используется, когда повреждений в ДНК становиться много.
• Степень индукции SOS-репарации пропорциональна количеству повреждений в ДНК.
• При небольшом числе повреждений увеличивается число репаративных белков.
• При большем числе повреждений приостанавливается деление клетки и индуцируется синтез еще большего числа репаративных белков.
• При еще большем числе повреждений в клетке синтезируются белки, которые способствуют ДНК-полимеразе осуществлять синтез дочерней цепи ДНК, используя в качестве матрицы дефектные звенья материнской цепи.
• В связи с этим в дочерней цепи ДНК появляется много ошибок.
• Благодаря SOS-репарации происходит удвоение ДНК и клетка может разделиться.
Дочерние клетки выживут, если жизненно важные функции, закодирован
• 18. Общие представления о транскрипции. В результате транскрипции происходит матричный синтез РНК, образуются иРНК, тРНК, рРНК и др. РНК, синтез РНК фермент РНК-полимераза. 3’-конца промотором – участком с которым связывается РНК-полимераза,со стороны 5’-конца – терминатором – участком, в котором прерывается синтез РНК.Последовательность ДНК, ограниченная промотором и терминатором, представляет собой единицу транскрипции – транскриптон. Выделяют три стадии: инициацию, элонгацию и терминацию. На стадии инициации РНК-полимераза, взаимодействуя с промотором, вызывает расхождение цепей ДНК и начинает синтез молекулы РНК. В ходе элонгации новосинтезированная цепь РНК образует короткие отрезки гибридной двойной спирали ДНК-РНК, которые необходимы для правильного считывания цепи ДНК. Как только РНК-полимераза достигнет терминирующих последовательностей, запускается последняя стадия – терминация. По ее завершению происходит освобождение вновь синтезированной цепи РНК).РНК-полимераза в качестве субстратов для синтеза РНК использует рибонуклеозид-5’-фосфаты (АТФ, ГТФ, ЦТФ и УТФ). РНК-полимераза удлиняет цепь, присоединяя нуклеотиды к 3’-концу, т.е. наращивает цепь в направлении 5’®3’.РНК-полимераза требует для своей работы в качестве матрицы одну из цепей ДНК.РНК-полимераза не нуждается в затравке.РНК-полимераза последовательно добавляет нуклеотиды к 3’ – концу полинуклеотидной цепи.Нуклеотиды в процессе транскрипции присоединяются к цепи в соответствии с принципом комплементарности. Т.о., образовавшаяся в результате транскрипции РНК комплементарна матричной цепи ДНК.
|
|
|
•
• 19. Транскрипция у прокариот. Прокариоты содержат одну РНК-полимеразу.Ее субъединичный состав - a2bb’s. s -субъединица необходима для узнавания промотора.После инициации s-субъединица отделяется.Элонгацию осуществляет комплекс a2bb’ (кор-фермент).s -субъединицы различаются в зависимости от того, какую группу промоторов должна узнавать РНК-полимераза. Как правило s-факторы узнаются блоки, отстоящие на 10 и 35 нуклеотидов от точки начала транскрипции. Существует два типа терминаторов: r-зависимые терминаторы и r-независимые терминаторы.r-Независимая терминация обеспечивается образованием РНК шпильки в процессе транскрипции и последующей олигоуридиловой последовательностью.Шпилька приводит к паузе в транскрипции, а олигоуридил-олигоадениловый дуплекс, как наименее стабильный, диссоциирует в ходе паузы.Инициация транскрипции может регулироваться в результате изменения количества и активности различных s-факторов в клетке.Таким способом регулируется метаболизм азота.При недостатке азота происходит фосфорилирование белка NtrC. фосфорилированный белок NtrC взаимодействует с комплексом РНК-полимеразы и s 54.Происходит активация транскрипции генов, необходимых для восстановления азотного баланса.
|
|
|
•
• 20. Транскрипция у эукариот.В клетках эукариот присутствуют три РНК-полимеразы.РНК-полимераза I транскрибирует гены рРНК 18S, 28S и 5,8S.РНК-полимераза II транскрибирует гены иРНК.РНК-полимераза III транскрибирует гены тРНК, 5S рРНК и мяРНК. РНК-полимераза I эукариот является большим ферментом, построенным по меньшей мере из 11 субъединиц, транскрибирует гены рРНК.Гены рРНК представлены в геноме клетки большим числом копий.Гены рРНК кодируют три наиболее крупных рРНК – 18S рРНК, 5,8 S и 28S рРНК.Рибосомные гены транскрибируются в ядрышках в виде единого 45S-РНК транскрипта (13000 н).45S-РНК расщепляется с образованием 28S-РНК (около 5000 нуклеотидов), 18S-РНК (около 2000 нуклеотидов) и 5,8S-РНК (около 160 нуклеотидов). 5S рРНК транскрибируется отдельно от других рРНК РНК-полимеразой III.Транскрипция генов рРНК соответствует 30- 40% от общего объема клеточной транскрипции.Гаплоидные клетки человека содержат около 200 генов рРНК, которые распределены в виде небольших кластеров по пяти хромосомам.Гены рРНК расположены в виде серии тандемных повторов, отделенных друг от друга спейсером.В конце спейсера находится сайт терминации (Т), он же отвечает и за реинициацию нового транскрипта.В спейсере имеются энхансеры (40-60 п.н.), активирующие транскрипцию в десятки раз. РНК-полимераза II, транскрибирует гены, которые затем будут транслированы в белки.РНК-полимераза II содержит более 10 субъединиц.Но самостоятельно она не в состоянии осуществлять транскрипцию.Чтобы РНК полимераза могла транскрибировать ген, требуется взаимодействие около 40 - 50 белковых молекул – факторов транскрипции.Альфа-аманитин обладает необычно сильным сродством к РНК-полимеразе II.РНК-полимераза I нечувствительна к нему, а РНК-полимераза III чувствительна слабо.Попадая в клетку, аманитин связывает этот фермент, что приводит к прекращению синтеза белков и к разрушению клетки (цитолизу).
|
|
|
• 21. Энхансеры и сайленсеры. Энхансеры и сайленсеры регулируют интенсивность экспрессии.генов.Энхансеры – регуляторные последовательности ДНК, с которыми взаимодействуют белки-активаторы, в результате транскрипция генов многократно усиливается.Сайленсеры – регуляторные последовательности ДНК, с которыми взаимодействуют белки-ингибиторы, в результате транскрипция генов ослабевает. В природе энхансеры могут занимать различное положение.Чаще перед геном за несколько сот п.н. и даже за несколько тысяч п.н.Энхансеры обнаружены в интронах.Энхансеры встречаются за геном.Свойствами энхансера могут обладать кодирующие последовательности.Энхансеры можно переносить от одного гена к другому.Один ген может иметь несколько энхансеров.Граница энхансера размыта – удаление нуклеотидных последовательностей постепенно снижает активность энхансера.Энхансеры могут содержать блоки, мутации в которых резко снижает их активность.Организация сайленсеров имеют много общего с организацией энхансеров.Энхансеры в зависимости от регуляторного фактора могут вести себя как сайленсоры.
|
|
|
