4.1 Сравнение аналитической и имитационной моделей. 17
4.2 Анализ экспериментальных данных. 19
Заключение. 20
Список литературы к реферату. 21
Список использованных источников. 21
Список публикаций соискателя. 22
Предметный указатель к реферату. 23
Интернет ресурсы в предметной области исследования. 24
Действующий личный сайт в WWW... 25
Граф научных интересов. 26
Презентация магистерской диссертации. 27
Тестовые вопросы.. 28
Список литературы к выпускной работе. 29
Приложения. 31
Список обозначений ко всей выпускной работе
Реагенты
AT М
g-актин в АТФ форме
AD М
g-актин в АДФ форме
ATF
f-актин в АТФ форме
ADF
f-актин в АДФ форме
FIB
Плюс-концы филамент
FTB
Плюс-концы с последним актином в АТФ форме
FDB
Плюс-концы с последним актином в АДФ форме
FIP
Минус-концы филамент
FTP
Минус-концы с последним актином в АТФ форме
FDP
Минус-концы с последним актином в АДФ форме
CBM
Блокирующий протеин плюс-конца в свободной форме
CBF
Блокирующий протеин плюс-конца в связанной форме
FOM
Формин в свободной форме
FOF
^
FxB
FxP
Формин в связанной форме
Позиция между двумя актинами в филаменте
Все типы плюс-концов филамент
Все типы минус-концов филамент
Реакции
SNUC
Нуклеация
FNUC
Формин-инициируемая нуклеация
ASTB
Элонгация плюс-конца частицей АТФ актина
ASDB
Элонгация плюс-конца частицей АДФ актина
ASTP
Элонгация минус-конца частицей АТФ актина
ASDP
Элонгация минус-конца частицей АДФ актина
ASTF
Элонгация плюс-конца с фомином частицей АТФ актина
DITB
Диссоциация АТФ актина с плюс-конца
DIDB
Диссоциация АДФ актина с плюс-конца
DITP
Диссоциация АТФ актина с минус-конца
DIDP
Диссоциация АДФ актина с минус-конца
ASCB
Блокировка плюс-конца
DICB
Разблокировка плюс-конца
CBNU
Нуклеация c участием блокирующего протеина
DIFB
Отсоединение формина от плюс-конца
ASFB
Присоединение формина к плюс-концу
TTOD
Переход f-актина из АТФ в АДФ форму
DTOT
Переход g-актина из АДФ в АТФ форму
Реферат на тему «Применение информационных технологии при моделировании биохимических реакции в процессах полимеризации актина»
Введение
В настоящее время вычислительный эксперимент является одним из основных инструментов развития таких естественнонаучных областей как физика, химия, биология. Как правило, исследуемые в этих научных дисциплинах системы характеризуются значительным числом взаимодействующих компонентов (в свою очередь являющихся подсистемами), многообразием связей между своими компонентами, нелинейностью поведения и, как следствие, сложностью прогнозирования. Поскольку, зачастую, нет простых формул, описывающих поведение модели, а стало быть, и объекта, который описывается моделью, то единственный путь – свести дело к вычислениям, применению численных методов для решения задач.
Одной из наиболее актуальных в настоящее время областей для применения является применение моделирования к молекулярным биологическим процессам и системам. Отличием биологических (клетки) и биомолекулярных (протеины, ДНК, и т.д.) объектов является их чрезвычайная сложность. В то же время, без детальной информации об этих объектах и клеточных процессах невозможна разработка лекарственных препаратов и эффективного лечения таких заболеваний, как различные типы рака, вирусные инфекции, возрастные заболевания (болезнь Альцгеймера, остеопороз).
Глава 1.Обзор литературы
Настоящая работа посвящена построению и анализу моделей полимеризации актина – важного клеточного процесса, обуславливающего динамику клеток. Изучение данного явления кроме научно-исследовательского имеет и прикладное значение. Некоторые патогенные бактерии используют полимеризацию актина для движения внутри инфицированной клетки [1]. Образование и динамика раковых клеток и метастаз в значительной степени зависят от полимеризации актина. Поэтому управление и контроль над данным процессом может позволить снизить активность патогенов и повысить стабильность здоровых клеток.
Несмотря на большое число теоретических работ посвященных этой тематике, формализм моделирования процессов актин-полимеризации не до конца разработан. Таким образом, актуальной является разработка формализма в рамках парарадигмы имитационного моделирования. Хотя подобные работы проводились ранее [2], [3], по-прежнему ощущается недостаток систематического рассмотрения вопроса моделирования актин-полимеризации. Модель Альбертса и Одела разрабатывалась в основном для анализа механических особенностей роста актин-филамент и уделяет очень мало внимания моделированию биохимических реакций. Модель представленная Карлссоном [4] использует крайне неэффективный алгоритм моделирования, и может быть значительно ускорена при использовании алгоритмов моделирования Гиллеспи.
Одним из важнейших вопросов, связанных с данной темой, является построение методики анализа экспериментальных данных с использованием имитационных моделей. Такой подход для обработки результатов биофизических экспериментов уже не нов и использовался при изучении сложных процессов и систем. Он ранее описан в работах групп Схафсмы и Хэмминги [5-7] для задач анализа данных флуоресценции.
Целью данной работы является разработка, реализация и тестирование алгоритмов имитационного моделирования полимеризации актина, позволяющих исследовать влияние различных реагентов и реакций на динамику роста филамент.
В рамках работы были изучены различные процессы, участвующие в полимеризации актина, включая: образование филамент, их элонгацию и диссоциацию мономеров актина на концах филамент, блокировку, фрагментацию, гидролиз, реакции с участием формина. Все разработанные алгоритмы и процессы были реализованы в виде програмного продукта, написанного на языке Java. При выборе языка программирования были учтены несколько очевидных преимуществ, обеспечиваемых Java: независимость от архитектуры компьютера, переносимость, а главное, кроссплатформенность, что позволяет использовать созданную программу на различных компьютерах.
Во второй главе описываются биофизические аспекты процессов актин-полимеризации. Детально рассмотрены основные реагенты и взаимосвязи между ними. Вторая глава работы также описывает методику моделирования химических реакций. Анализируются достоинства и недостатки различных методов моделирования с точки зрения их точности, быстроты вычислений, времени необходимого на разработку конечного алгоритма, применимости для особых случаев.
В третьей главе рассказывается о разработанной модели и ее реализации, обсуждаются сделанные допущения.
Заключительная глава посвящена тестированию алгоритмов и анализу результатов моделирования. Влияние всех учтенных в модели контролирующих механизмов изучено, полученные результаты обоснованы. Также, было проанализировано быстродействие разработанных имитационных алгоритмов. Результаты, полученные с помощью разработанной имитационной модели, были сравнены с аналитическим решением, а также, с экспериментальными данными.
Глава 2.Методика исследований
2.1 Актуальность изучения процессов полимеризации актина
Актин является белком, входящим в состав клеточного скелета, который может находиться в одной из двух взаимопереходящих форм — глобулярной и фибриллярной. Полимеризацией и называется способность глобулярной формы актина (далее g-актин) организовываться в длинные цепи актинов (далее f-актин), которые называются филаментами. Данный процесс играет важную роль в процессах миграции, деления и объединения клеток. Поэтому не удивительно, что нарушение клеточного скелета напрямую связано с такими болезнями как рак, миопарез и нейродегенеративные заболевания [8]. Хотя многочисленные исследования проводятся в настоящее время, пока не существует полного физико-химического описания происходящих процессов. До сих пор еще остается неизвестно, почему рост филаментов происходит достаточно быстро по сравнению с другими процессами, почему удлинение филаментов способно воздействовать на мембрану, вызывая движение клеток в определенном направлении, каков механизм передачи энергии АТФ для роста филаментов, и как можно управлять этими процессами [9].
В данной работе исследуются вопросы, связанные со стохастическим моделированием и анализом процессов полимеризации актина. Поскольку проведение экспериментов с реальными организмами часто не дает возможности получить полной картины процесса, что обуславливает использование имитационных моделей, например, для нахождения скоростей реакции.
2.2 Реагенты и реакции при полимеризации актина
Мономеры актина имеют грушевидную форму, и при полимеризации возникает спирально закрученная полярная нить с различающимися концами: заостренным (минус) и оперенным (плюс) концом (рис.1).
Такие названия появились в связи с тем, что при взаимодействии актиновых нитей с фрагментами молекулы моторного белка миозина образуется комплекс, имеющий под электронным микроскопом стекловидную форму. При этом острые стрелы указывают на заостренный конец, а ее оперение обращено в сторону противоположного конца филамента. Полимеризация актина происходит в две стадии. Первая стадия
Рисунок.1-Полярная нить актина и круговорот мономеров при полимеризации актина. Рост нити происходит за счет активного присоединения мономеров к оперенному (плюс) концу нити и их более медленной диссоциации с заостренного (минус) конца. По[10]
носит название “нуклеация”, то есть создание ядра (nucleus) или затравки из первых трех мономеров актина. Димер (комплекс двух мономеров) является нестабильной структурой и легко разрушается. Именно нуклеация определяет общую скорость полимеризации. Вторая стадия, удлинение нити, протекает легче, с большей скоростью.
В пробирке мономерный актин может присоединяться и диссоциировать с обоих концов нити, но присоединение происходит быстрее к плюс-концу. Процесс непрерывного присоединения мономеров к оперенному (плюс) концу и их диссоцации с заостренного (минус) представляет собой непрерывный круговорот мономеров. В результате теплого движения филаментов происходит процесс спонтанной фрагментации - самопроизвольный распад цепочки f-актина на две части.
В живой клетке процесс полимеризации актина может быть не похож на такой круговорот мономеров в пробирке, поскольку полярные концы актинового филамента могут быть несвободными.
В клетке присутствуют десятки так называемых актин связывающихся белков, которые сильно влияют на процесс полимеризации актина. Имеется четыре места действия белков: с мономером актина, с плюс-концом, с минус-концом, с боковой поверхностью. Некоторые из них могут блокировать активно растущий оперенный плюс- конец, так называемые кэп-белки, или блокирующие протеины (от англ. cap – шапочка), прекращая таким образом полимеризацию уже существующих нитей и освобождая мономерный актин для построения новых нитей. Другие белки могут разрезать нити актина, формируя тем самым новые фрагменты с заостренными и оперенными концами. Существуют белки, связывающие мономеры актина и таким образом делающие их недоступными для полимеризации (например, профилин), белки, деполимеризующие актин и т.п.[10].
Как f-актин, так и g-актин могут присутствовать в системе в двух состояниях: АТФ-состояние соответствует повышенной вероятности процессов полимеризации, актин в АДФ состоянии быстрее диссоциирует. Переход из АТФ в АДФ состояние происходит, когда молекула актина находится в составе филамента. Обратный процесс возможен для актина в g-форме с участием профилина. Изучение влияния данного протеина достаточно важно, поскольку профилин предположительно является источником энергии для процессов полимеризации. Далее остановимся на некоторых дополнительные протеинах, влияющих на протекающие процессы более подробно:
• АДФ/кофилин – протеин, который может присоединяться к f-актину и стимулировать его отделение от филамента. В таком случае происходит диссоциация мономера, находящегося в любом месте исходной филаменты с ее разделением и образованием двух новых. С одной стороны это производит разрушающий эффект, с другой стороны образование новых концов может ускорять элонгацию при достаточно большой концентрации g-актина
• Кэп-белки (блокирующие протеины) могут присоединяться к свободным концам филаментов, «закрывая» их, то есть, останавливая процессы элонгации и диссоциации для данного конца. Существуют различные виды блокирующих протеинов, которые присоединяются к строго отпределенным концам филаментов (плюс- или минус- концам, оказывая тем самым качественно различное влияние на рост филаментов.
• ARP2/3–комплекс актино-подобных протеинов, имеющих повышенное сродство к минус концам филаментов и может служить матрицей,на которой происходит образование новых актиновых нитей(нуклеация)(рис.2.а).Кроме того, комплекс может присоединяться к боковым сторонам нитей и таким образом обеспечивать их ветвление(рис.2.б).Эффект ветвления вызывает как правило ускорение элонгации и сильно влияет на механические свойства филамент, которые рассматриваются при исследовании динамических явлений, возникающих в процессе полимеризации актина. Интересно,что боковые нити актина отходят от основной нити строго под углом 70°,образуя жесткую сеть[10].
• Формин является протеином, ускоряющим нуклеацию филаментов. В процессе роста формин остается прикрепленным к плюс-концу, ускоряя также реакцию элонгации [11].
2.3 Стохастический подход к моделированию биохимических реакций
Имитационное моделирование с использованием парадигмы Монте-Карло основано на получении большого числа реализаций стохастического (случайного) процесса. Данный процесс формируется таким образом, чтобы его вероятностные характеристики совпадали с аналогичными величинами решаемой задачи.
Введем как функцию, описывающую количество молекул iого вещества в момент времени t. Тогда в момент времени что система находится в состоянии . Чтоб описать изменение ее состояния с течением времени нужно ответить на вопросы: когда наступит следующая реакция и что это будет за реакция. Однако из-за стохастичности реакций эти ответы могут быть даны только с определенной вероятностью. Учитывая предыдущие утверждения, введем функцию , такую, что выражение:
(1)
- определяет вероятность того, что на интервале времени в системе произойдет реакция в объеме V, если известно, что в момент времени она находилась в состоянии .
называется функцией плотности вероятности реакции, поскольку, с математической точки зрения, это объединенная функция плотности вероятности по непрерывному параметру и дискретному . Ценность значений переменных и в том, что они дают ответ на те два вопроса, которые были заданы выше: характеризует промежуток времени, за который произошла реакция, а величина непосредственно указывает на номер реакции, которая имела место. Первым шагом на пути поиска приемлемого метода для присваивания численных значений для и является получение аналитического выражения для .
Для каждой реакции определим функцию как количество комбинаций отдельных реагентов , которые возможны в состоянии .
Тогда если имеет вид:
продукт реакции, получим:
; (2)
если имеет вид: продукт реакции, тогда
(3)
Таким образом функция представляет собой число всевозможных комбинаций молекул, принимающих участие в реакции μ из набора .Отсюда:
- вероятность того, что на интервале времени с системой произойдет реакция в объеме V, если известно, что в момент времени она находилась в состоянии , где - стохастическая скорость реакции.
Тогда можно вычислить вероятность (1) как произведение(4).
(вероятность того, что интервале времени с системой не произойдет никакой реакций в объеме V, если известно, что в момент времени она находилась в состоянии ); -последующая вероятность, что реакция произойдет в интервале :
(4)
Для того, чтоб найти выражение для отметим, что -вероятность того, что за промежуток с системой в состоянии не произойдет никакой реакции. Таким образом:
(5),
откуда легко получается, что
(6)
Подставляя (6) в (4) получим, что функция плотности вероятности реакции, описанная в (1) равна
, (7)
где .
Отметим, что в (7) - это строгое математическое следствие фундаментальной гипотезы, которое зависит от всех постоянных реакции (не только от )и от текущего значения молекул всех реагирующих веществ(а не только от реагентов )[21].
Для отдельной реализации случайного процесса в каждый момент времени система находится в определенном состоянии. Из этого состояния системы выбирается переход в другое состояние системы на основе вероятностей всех возможных переходов. Ключевым является выбор случайных чисел, используя машинный генератор случайных чисел и их использование для выбора превращений.
Первый из методов, предложенных Гиллеспи, получил название «прямой метод»[12]. Он однозначно вычисляет какая реакция произойдет следующей и когда она произойдет. Второй предложенный метод Гиллеспи назвал «методом первой реакции»[13].Суть метода заключается в генерации для каждой из реакций предполагаемого времени наступления реакции (τ),затем выбирается реакция с наименьшим τ (“первая реакция”)и выполняется данная реакция (μ) в момент времени τ. Позднее был предложен метод τ-leap [14], представляющий собой метод с постоянным шагом по времени. Используя пуассоновскую аппроксимацию для потока реакций, он позволяет оптимизировать время моделирования. Существует широкий спектр задач, где метод τ-leap очень хорошо аппроксимирует стохастичное поведение модели,однако он является неприменимым для жестких стохастичных задач.
2.4.1 Прямой метод
Этот метод основан на том, что каждая из двумерных функций плотности вероятности может быть записана как произведение двух одномерных функций плотностей, данная операция известна как операция «условия». Введем условную вероятность как произведение
(8)
где –вероятность того, что следующая реакция произойдет в интервале времени от до ,а -вероятность того, что следующей реакцией будет реакция , при условии, что она происходит в момент времени .
Поскольку
(9),
где -количество комбинаций молекул отдельных реагентов для реакции , обнаруженных в объеме V в момент времени t, заменяя на (9) приходим к результату:
(10)
(11),
где для удобства заменим
, .
Вероятность зафиксировать какую-нибудь реакцию в течение короткого времени определяется выражением:
(12)
В данном выражении - вероятность зафиксировать хотя бы одну реакцию за единицу времени; - число всевозможных реакций в системе. Величина определяет вероятность того, что ни одна реакция не произойдет за время .
Видно, что в данном случае не зависит от .Также мы должны учесть, что эти одновариантные функций плотностей нормированы в своей области определения:
(13)
(14)
Идеей прямого метода является генерация произвольного значения τ, соответствующему (13), а затем генерация произвольного целого μ,соответствующего в (14). Результатом является произвольная пара ,помещенная в .
Таким образом, время наступления первой реакции рассчитывается методом обратной функции по формуле (15):
(15) (16)
Номер реакции определяется методом суперпозиции по формуле (16):
(16)
где - независимые реализации базовой случайной величины
Рисунок 2- Алгоритм прямого метода Гиллеспи
Схема алгоритма моделирования представлен на рисунке 2. Охарактеризуем каждый из блоков:
1. Задание начальных концентраций,скоростей и механизмов реакций, моделируемого объема, времени моделирования, и установление .
2. Вычисление для всех и .
3. Вычисление времени наступления реакции, используя по формуле (15).
4. Генерирование случайного , определяющего номер произошедшей реакции. Определение номера произошедшей реакции по формуле (16).
5. Замена количества молекул согласно произошедшей реакции, устанавливаем .
6. Переход к шагу 2.
Как уже писалось, этот алгоритм использует два случайных числа за итерацию, что занимает времени, пропорционально количеству реакций, для пересчета и μ [12].
2.4.2 Метод первой реакции
Вместе с первым методом Гиллеспи был разработан алгоритм «первой реакции»,который генерирует предполагаемое время наступления каждой из реакции,- время, когда наступит данная реакция, если никакая другая не наступит раньше, потом полагается, что μ -реакция с наименьшим предполагаемым временем, τ переобозначается как .
Алгоритм:
1.Задание начальных концентраций,реакций и их скоростей, установление ;
2.Вычисление для всех i.
3. Генерирование для каждого i с помощью метода обратных функций возможного времени ,
4.Пусть μ-реакция с наименьшим временем ¸
5.Полагаем τ= ¸.
6.Меняем количество молекул согласно произошедшей реакции μ,устанавливаем ;
7.Переход к шагу 2.
На первый взгляд, эти два метода кажутся очень разными, но они эквивалентны, так как распределения вероятности, используемые для вычисления τ и μ одинаковы. Как уже писалось, данный алгоритм также использует r случайных чисел за итерацию (r- количество реакций), что затратит время, пропорционально r, чтоб пересчитать все и столько же, чтоб вычислить наименьшее [15].




 как функцию, описывающую количество молекул iого вещества в момент времени t. Тогда в момент времени
как функцию, описывающую количество молекул iого вещества в момент времени t. Тогда в момент времени  что система находится в состоянии
что система находится в состоянии  . Чтоб описать изменение ее состояния с течением времени нужно ответить на вопросы: когда наступит следующая реакция и что это будет за реакция. Однако из-за стохастичности реакций эти ответы могут быть даны только с определенной вероятностью. Учитывая предыдущие утверждения, введем функцию
. Чтоб описать изменение ее состояния с течением времени нужно ответить на вопросы: когда наступит следующая реакция и что это будет за реакция. Однако из-за стохастичности реакций эти ответы могут быть даны только с определенной вероятностью. Учитывая предыдущие утверждения, введем функцию  , такую, что выражение:
, такую, что выражение: (1)
(1)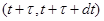 в системе произойдет реакция
в системе произойдет реакция  в объеме V, если известно, что в момент времени
в объеме V, если известно, что в момент времени  называется функцией плотности вероятности реакции, поскольку, с математической точки зрения, это объединенная функция плотности вероятности по непрерывному параметру
называется функцией плотности вероятности реакции, поскольку, с математической точки зрения, это объединенная функция плотности вероятности по непрерывному параметру 
 и дискретному
и дискретному 
 . Ценность значений переменных
. Ценность значений переменных  в том, что они дают ответ на те два вопроса, которые были заданы выше:
в том, что они дают ответ на те два вопроса, которые были заданы выше: 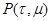 .
. как количество комбинаций отдельных реагентов
как количество комбинаций отдельных реагентов  , которые возможны в состоянии
, которые возможны в состоянии  имеет вид:
имеет вид: продукт реакции, получим:
продукт реакции, получим: ; (2)
; (2) продукт реакции, тогда
продукт реакции, тогда (3)
(3) представляет собой число всевозможных комбинаций молекул, принимающих участие в реакции μ из набора
представляет собой число всевозможных комбинаций молекул, принимающих участие в реакции μ из набора  .Отсюда:
.Отсюда: - вероятность того, что на интервале времени
- вероятность того, что на интервале времени  с системой произойдет реакция
с системой произойдет реакция  она находилась в состоянии
она находилась в состоянии  , где
, где  - стохастическая скорость реакции.
- стохастическая скорость реакции. (вероятность того, что интервале времени
(вероятность того, что интервале времени  с системой не произойдет никакой реакций в объеме V, если известно, что в момент времени
с системой не произойдет никакой реакций в объеме V, если известно, что в момент времени  -последующая вероятность, что реакция
-последующая вероятность, что реакция  произойдет в интервале
произойдет в интервале  :
: (4)
(4) -вероятность того, что за промежуток
-вероятность того, что за промежуток  с системой в состоянии
с системой в состоянии  (5),
(5), (6)
(6)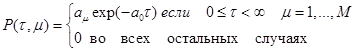 , (7)
, (7) .
. в (7) - это строгое математическое следствие фундаментальной гипотезы, которое зависит от всех постоянных реакции (не только от
в (7) - это строгое математическое следствие фундаментальной гипотезы, которое зависит от всех постоянных реакции (не только от  )и от текущего значения молекул всех реагирующих веществ(а не только от реагентов
)и от текущего значения молекул всех реагирующих веществ(а не только от реагентов  )[21].
)[21]. как произведение
как произведение  (8)
(8) –вероятность того, что следующая реакция произойдет в интервале времени от
–вероятность того, что следующая реакция произойдет в интервале времени от  до
до  ,а
,а  -вероятность того, что следующей реакцией будет реакция
-вероятность того, что следующей реакцией будет реакция  (9),
(9), -количество комбинаций молекул отдельных реагентов для реакции
-количество комбинаций молекул отдельных реагентов для реакции  , обнаруженных в объеме V в момент времени t, заменяя
, обнаруженных в объеме V в момент времени t, заменяя  на (9) приходим к результату:
на (9) приходим к результату:
 (10)
(10)
 (11),
(11),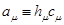
 ,
, 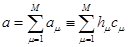 .
.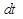 определяется выражением:
определяется выражением:
 - вероятность зафиксировать хотя бы одну реакцию за единицу времени;
- вероятность зафиксировать хотя бы одну реакцию за единицу времени;  - число всевозможных реакций в системе. Величина
- число всевозможных реакций в системе. Величина  определяет вероятность того, что ни одна реакция не произойдет за время
определяет вероятность того, что ни одна реакция не произойдет за время  .
. не зависит от
не зависит от 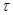 .Также мы должны учесть, что эти одновариантные функций плотностей нормированы в своей области определения:
.Также мы должны учесть, что эти одновариантные функций плотностей нормированы в своей области определения: (13)
(13) (14)
(14)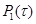 (13), а затем генерация произвольного целого μ,соответствующего
(13), а затем генерация произвольного целого μ,соответствующего  ,помещенная в
,помещенная в  (15) (16)
(15) (16)
 определяется методом суперпозиции по формуле (16):
определяется методом суперпозиции по формуле (16): (16)
(16)
 - независимые реализации базовой случайной величины
- независимые реализации базовой случайной величины
 .
. для всех и
для всех и  по формуле (15).
по формуле (15).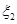 , определяющего номер произошедшей реакции. Определение номера произошедшей реакции по формуле (16).
, определяющего номер произошедшей реакции. Определение номера произошедшей реакции по формуле (16). .
. .
. ;
; ,
, и столько же, чтоб вычислить наименьшее
и столько же, чтоб вычислить наименьшее 