 |


Физиологическое действие награды
|
|
|
|
Рассмотренные выше темы оптимального уровня возбуждения и процесса противодействия привлекают наше внимание к вопросу о том, что общего могут иметь различные мотивы с точки зрения физиологии. Все они включают некоторый оптимальный уровень возбуждения (даже если этот оптимум меняется от мотива к мотиву); все, вероятно, имеют свой процесс противодействия, поддерживающий стабильность организма. Но существуют ли какие-то общие физиологические черты, лежащие в основе различ-
 1 См.: Solomon R.L., Corbit J.D. An opponent-process theory of motivation: I. Temporal
1 См.: Solomon R.L., Corbit J.D. An opponent-process theory of motivation: I. Temporal
dynamics of affect // Psychological Review. 1974. Vol. 81. P. 119-145; Solomon R.L. The
opponent-process theory of acquired motivation: The costs of pleasure and the benefits of
pain // American Psychologist. 1980. Vol. 35. P. 691-712.
2 См.: Solomon R.L. The opponent-process theory of acquired motivation: The costs of
pleasure and the benefits of pain // American Psychologist. 1980. Vol. 35. P. 691-712.
Глейтман Г., Фридлунд А, РайсбергД. Мотивация 109
 ных мотивов? Может быть, есть некоторая отдельная область мозга, активация которой вызывает чувство, называемое нами «удовольствием» — будь то удовольствие от секса, прослушивания музыки или вкусной еды? Исследователи пытались ответить на этот вопрос, изучая действие награды с помощью электрической стимуляции различных зон головного мозга.
ных мотивов? Может быть, есть некоторая отдельная область мозга, активация которой вызывает чувство, называемое нами «удовольствием» — будь то удовольствие от секса, прослушивания музыки или вкусной еды? Исследователи пытались ответить на этот вопрос, изучая действие награды с помощью электрической стимуляции различных зон головного мозга.
Общие исследования были начаты в 1954 г., когда Джеймс Олдс и Питер Милнер обнаружили, что крысы начинают нажимать на педаль, если получают в награду один лишь короткий разряд электрического тока в определенную зону гипоталамуса и лимбической системы1. Подобное действие самостимуляции как награды было продемонстрировано на множестве примеров других животных, включая кошек, дельфинов, обезьян, а также и людей. Чтобы получить такую стимуляцию, крысы давили на педаль с частотой до 7000 нажатий в час в течение нескольких часов. Если им предлагали выбирать между едой и удовольствием, то крысы часто выбирали второе, даже если они буквально умирали от голода2.
|
|
|
Специфические и генерализованные центры удовольствия
Чем можно объяснить действие самостимуляции, подобное получению награды? Единственное возможное объяснение заключается в том, что стимуляция мозга вызывает ощущение, схожее с тем, которое оказывает сама награда. Так, стимуляция одной зоны может ввести мозг в заблуждение, что была получена настоящая еда, стимуляция другой зоны — что произошел сексуальный контакт, и т.д. Другой взгляд на проблему заключается в том, что самостимуляция вызывает более генерализованный, неспецифический вид удовольствия, подобно тому, которое дает получение любой награды. Еда, питье и секс, безусловно, различны между собой, но возможно, что та разная информация, которую они посылают в мозг (только что поел, попил, совершил половой акт), в конечном итоге сливается в единую функциональную систему, которая на все эти сигналы реагирует практически однотипным образом («чувствую себя хорошо»).
Большинство полученных данных говорят в пользу первого предположения — стимуляция разных зон мозга дает ощущение получения новой неспецифической награды. Например, животные выполняют определенные действия, чтобы получить электрический стимул в латеральную часть гипоталамуса (которая отвечает за чувство голода), но то, насколько активно они работают для получения этой награды, зависит от степени голода. Если они долго не ели, то начинают работать активнее, чем в другом
 1 См.: Olds J., Milner P. Positive reinforcement produced by electrical stimulation of
1 См.: Olds J., Milner P. Positive reinforcement produced by electrical stimulation of
septal areas and other regions of rat brains // Journal of Comparative and Physiological
Psychology. 1954. Vol. 47. P. 419-427.
2 См.: Spies G. Food versus intracranial self-stimulation reinforcement in food deprived
rats // Journal of Comparative and Physiological Psychology. 1965. Vol. 60. P. 153-157.
110 Тема 12. Психология мотивации
|
|
|
 случае. Это значит, что мозг рассматривает стимуляцию этой зоны как эквивалент получения пищи. Аналогичный эффект наблюдается при стимуляции зон, связанных с жаждой и сексуальным поведением1.
случае. Это значит, что мозг рассматривает стимуляцию этой зоны как эквивалент получения пищи. Аналогичный эффект наблюдается при стимуляции зон, связанных с жаждой и сексуальным поведением1.
Допаминовая теория удовольствия и наркотического воздействия
Что является физиологическим базисом самостимуляции? Многие исследователи считают, что ответ содержится в допаминовых каналах головного мозга. Известно, что самостимуляция наиболее эффективна, когда она направлена на сгусток нервных волокон, который называется медиальным пучком переднего мозга. Эти волокна тянутся от области среднего мозга к гипоталамусу и, по мнению некоторых исследователей, вызывают активность других клеток. Эти волокна проходят от одной части среднего мозга (вентромедиальная область — ВМО) к зоне в области переднего мозга, называемой передней комиссурой <...>. Нейромедиатором в этих каналах является допамин. Это доказывает тот факт, что если препараты, являющиеся антагонистами допамина, вводятся в близлежащие области мозга, стимуляция действует намного слабее2.
Многие ученые объясняют эти явления тем, что активация нервных волокон, исходящих из передней комиссуры, воспринимается мозгом как неврологический эквивалент: «это хорошо — давай еще раз!». Это, вероятно, может объяснить некоторые явления наркотической зависимости. И кокаин, и амфетамины повышают уровень допамина в синапсе. Кокаин блокирует возврат допамина в проводящий канал (благодаря чему он остается в этом канале дольше); амфетамины действуют так же, а кроме того, повышают выделение допамина в аксоновом окончании. Попадая в кровь, оба наркотика скоро находят область передней комиссуры и запускают по нервным путям механизм «это хорошо — давай еще раз!». Есть еще очень много непонятного в данной области. Очевидно, что допамин играет решающую роль в физиологическом механизме награды, и мы можем выделить некоторые структуры мозга, имеющие для этого механизма большое значение. Однако мы до сих пор не можем просуммировать все разнообразные знания относительно мозговой стимуляции, чтобы создать единую и непротиворечивую картину биологического базиса для механизма действия награды или, даже больше, биологической основы удовольствия. Когда мы сможем это сделать, то поймем не только то, что крысы (а также собаки и дельфины) нажимают на
|
|
|
 1 См.: Olds M.E., Fobes T. The central basis of motivation: Intracranial self-stimulation
1 См.: Olds M.E., Fobes T. The central basis of motivation: Intracranial self-stimulation
// Annual Review of Psychology. 1981. Vol. 32. P. 523-574.
2 См.: Gallistel C.R., Shizgal P., Yeomans J. A portrait of the substrate for self-stimulation
// Psychological Review. 1981. Vol. 88. P. 228-273; Stellar J.R, Stellar E. The Neurobiology
of Motivation and Reward. N.Y.: Springer Verlag, 1985; Wise RA.. Rompre P.P. Brain dopamine
and reward // Annual Review of Psychology. 1989. Vol. 40. P. 191-226.
Глейтман Г., Фридлунд А., РайсбергА- Мотивация 111
 педаль, желая получить небольшой разряд электрического тока в определенную зону головного мозга. Мы сможем понять работу неврологического механизма, лежащего в основе действия естественных наград и мотивов, а также понять, что именно в мозгу заставляет людей и животных отмечать некоторые события как события, которые они хотят пережить вновь (например, поесть, когда голоден; совершить половой акт, когда испытываешь сексуальное возбуждение; и т.д.). И мы, возможно, поймем также некоторые биологические механизмы, лежащие в основе «неестественных» видов награды, таких, как наркотики, употребляя которые, множество людей становятся зависимыми от них.
педаль, желая получить небольшой разряд электрического тока в определенную зону головного мозга. Мы сможем понять работу неврологического механизма, лежащего в основе действия естественных наград и мотивов, а также понять, что именно в мозгу заставляет людей и животных отмечать некоторые события как события, которые они хотят пережить вновь (например, поесть, когда голоден; совершить половой акт, когда испытываешь сексуальное возбуждение; и т.д.). И мы, возможно, поймем также некоторые биологические механизмы, лежащие в основе «неестественных» видов награды, таких, как наркотики, употребляя которые, множество людей становятся зависимыми от них.
Природа мотивов
В течение последних десятилетий произошел огромный прогресс в области знаний о психологии и физиологии биологических потребностей и мотивов. Эти мотивы чрезвычайно разнообразны.
Некоторые из них: терморегуляция, голод и жажда — включают механизмы обратной связи; другие, такие, как потребность во сне или в сексе, их не имеют. Для некоторых мотивов, таких, как потребность в безопасности, оптимальный уровень довольно высок; для других, таких, как сон, — низок. Мы выделили психологические и физиологические черты, которые могут объединять мотивы, но в результате разнородность мотивов сохраняется. Возможно, наш поиск элементарного и общего не имеет смысла. Как и все ученые, психологи были бы счастливы получить ясные и точные определения. Но Природа создавала живой мир не для того, чтобы сделать психологов счастливыми; она создавала его, чтобы животные делали то, что они делают ради сохранения жизни и продолжения своих генов. Итак, в этой главе рассматривались мотивы, в основном формируемые внутренними механизмами. Эти мотивы связаны с физиологическими потребностями, которые мы должны удовлетворять, чтобы выжить. Но как только мы начинаем рассматривать мотивы в деталях, мы обнаруживаем, что наше мотивированное поведение зависит не только от физиологии, но и от наших знаний о той среде, в которой мы действуем. Например, то, что мы пьем, призвано не только сохранить наш водный баланс, но касается также и наших предпочтений, а в некоторых случаях, наших антипатий, привитых культурой. Большинству людей было бы очень трудно выпить стакан своей слюны, пусть даже она только что покинула их собственный рот; большинство из нас не смогли бы есть кузнечиков. То же касается и других мотивов — тех, которые направляют наше любопытство, наш страх, наше сексуальное возбуждение. Во всех этих случаях наше поведение строится на биологической основе, которая, подобно ткачу, вплетает нашу генетическую нить в ту ткань, которой в конечном счете и являемся мы сами.
|
|
|
Ж. Нюттен
[ПОЗНАВАТЕЛЬНАЯ И СОЦИАЛЬНАЯ
МОТИВАЦИИ]1
Познавательная мотивация
Вудвортс привлек внимание к потребности, о которой прежде мало говорили: потребности в восприятии (will to perceive). Он справедливо заметил, что наши отношения со средой определяются такими факторами, как желание «видеть, слышать, отчетливо видеть, отчетливо слышать, выяснить или понять то, что мы видим или слышим в данный момент»2. Аналогичные взгляды высказал Ниссен применительно к животным. Он отмечал у животного «почти постоянную активность, направленную на то, чтобы быть осведомленным об окружающей обстановке. Значительная часть повседневной активности грызуна, собаки, мартышки или шимпанзе состоит из движений тела и органов чувств, предназначенных для того, чтобы поддерживать контакт животного с происходящим вокруг»3. В начале века Мак-Дауголл (1908) говорил, что «у обезьян, животных, наиболее близких к человеку, явно выражено любопытство (или потребность в знании) и что оно не только заставляет животное приближаться к объекту и внимательно наблюдать за ним, но также и манипулировать им»4. Начиная с 1950 г. эти выводы были подтверждены и детально проанализированы в многочисленных экспериментах. <...>
 1 Экспериментальная психология / Под ред. П. Фресса, Ж. Пиаже. М.: Прогресс,
1 Экспериментальная психология / Под ред. П. Фресса, Ж. Пиаже. М.: Прогресс,
1975. Вып. V. С. 49-63.
2 Woodworth R.S. Reinforcement of perception // Amer. J. Psychol. 1947. 60. P. 119-124.
3 Ниссен Г.В. Филогенетическое сравнение // Стивене С.С. Экспериментальная
психология. М., 1960. Т. 1. С. 471.
|
|
|
4 Цит. по: McDougall W. An introduction to social psychology. L.: Methuen & Co.,
1948. P. 49-50.
Нюттен Ж, [Познавательная и социальная мотивации]
 1. Любопытство и новые объекты
1. Любопытство и новые объекты
А) В своем первом исследовании Берлайн1 помещал крыс в пустую экспериментальную клетку и позволял им знакомиться с ней в течение двух часов. Когда позднее крысы снова оказывались в этой клетке, они обнаруживали в ней три одинаковых предмета (первая группа из трех крыс — три кубика, вторая — три цилиндра), которые они могли обнюхивать два раза по 5 мин. Основная фаза эксперимента состояла в том, что, когда крыс в очередной раз помещали в клетку, один из трех знакомых предметов заменялся новым (два кубика и цилиндр — для первой группы, два цилиндра и кубик — для второй группы). Время, затраченное каждой крысой на исследование (обнюхивание) каждого из трех предметов, регистрировалось с помощью хронографа с тремя циферблатами (экспериментатор нажимал на соответствующую кнопку во время исследования каждого предмета). Время, затрачиваемое на обследование предметов каждой из 6 крыс экспериментальной группы, с точностью до 1 с приводится в первой строке таблицы 1. Различие между временем,
Таблица 1
Время (в секундах), затраченное на изучение каждого из трех предметов набора, один из которых является новым2
| Крысы I— VI | Группа А: 2 кубика и 1 цилиндр | Группа Б: 2 цилиндра и 1 кубик | ||||
| I | II | III | IV | V | VI | |
| Экспериментальная группа Контрольная группа | 8—7—2 2—0—0 | 0—4—1 7—12—9 | 0—2—1 4—7—7 | 4—1—3 2—3—2 | 1—3—1 7—2—6 | 1—1—2 3—8—2 |
Порядок цифр соответствует порядку, в котором появлялись предметы в клетке: выделенная цифра относится к новому предмету. 6 крыс экспериментальной группы 20 с изучали 2 знакомых предмета (т.е. в среднем 10 с на предмет). Время, затраченное на новый предмет, равно 22 с (т.е. на 12 с больше для 6 крыс или на 2 с для одного животного). Такое различие является статистически значимым (t = 3,55; p = 0,017). У контрольной группы такого различия не наблюдалось (средняя разность для одного животного - 0,17). Среднее время исследования 2 сходных (и уже знакомых) предметов в экспериментальной группе также значимо меньше, чем время исследования сходных (но не знакомых) предметов в контрольной группе (t = 2,27; р = 0,046).
1 См.: Berlyne D.E. Novelty and curiosity as determinants of exploratory behavior //
Brit. J. Psychol. 1950. 41. P. 68-80.
2 Там же. Р. 77-78.
8 Зак. 1662
114 Тема 12. Психология мотивации
 затраченным на исследование знакомого и нового предмета, оказывается статистически значимым. Контрольной группе, также состоявшей из шести крыс, после ознакомления с пустой клеткой сразу же предлагался набор разнородных предметов (2 кубика и цилиндр — первой группе, 2 цилиндра и кубик — второй). В контрольной группе не было различия между знакомыми и новыми предметами, ибо каждый из трех предметов исследовался в первый раз. Время, затраченное на исследование каждого предмета, показано во второй строке таблицы 1. Значимого различия между временем исследования в этом случае не наблюдается.
затраченным на исследование знакомого и нового предмета, оказывается статистически значимым. Контрольной группе, также состоявшей из шести крыс, после ознакомления с пустой клеткой сразу же предлагался набор разнородных предметов (2 кубика и цилиндр — первой группе, 2 цилиндра и кубик — второй). В контрольной группе не было различия между знакомыми и новыми предметами, ибо каждый из трех предметов исследовался в первый раз. Время, затраченное на исследование каждого предмета, показано во второй строке таблицы 1. Значимого различия между временем исследования в этом случае не наблюдается.
Б) Берлайну1 принадлежит и первая попытка экспериментального изучения «эпистемической» любознательности человека (в отличие от любопытства на уровне восприятия). Экспериментальной группе из 34 испытуемых — студентов университета — он предлагал список из 48 вопросов, касающихся 12 животных (беспозвоночных), 8 из которых были заведомо известны испытуемому, а 4 являлись достаточно экзотическими животными. Испытуемые должны были осуществить несколько операций с этими вопросами; они должны были выделить 12 вопросов, ответы на которые представляли для них наибольший интерес, указать вопросы, вызывавшие удивление, и пр. Затем им предлагался перечень высказываний, содержащих, между прочим, ответы на вопросы первого списка; после этого испытуемые получали последний список, в котором повторялись вопросы первого списка, но сформулированы они были на этот раз в виде неоконченных предложений, закончить которые должен был сам испытуемый, исходя из того, что стало ему известно из предыдущего списка (изложения фактов). Контрольной группе (состоявшей из такого же числа студентов университета) первый вопросник не предлагался, и они, следовательно, не осуществляли связанные с ним операции. После того как в списке животных они отмечали известных и неизвестных им животных, им предлагался перечень высказываний (таким образом были созданы условия, при которых любознательность испытуемых не была стимулирована с помощью предварительных вопросов). Наконец, испытуемые получали последний список предложений-вопросов, которые они должны были закончить. Эффект любознательности измерялся с помощью различных тестов на запоминание и интроспективных отчетов испытуемых. Было отмечено, в частности, что на те вопросы последнего теста, которые вызвали наибольший интерес испытуемых экспериментальной группы, были даны самые обстоятельные ответы. Точно так же лучше всего запоминались те изложенные факты (второй список), которые испытуемые считали ответами на вопросы первого списка. Наконец, столь же хорошо запоминались и данные, вызывавшие удивление.
1 См.: Berlyne D.E. An experimental study of human curiosity // Brit. J. Psychol. 1954. 45. P. 256-265.
Нюпен Ж. [Познавательная и социальная мотивации]
 2. Тенденция к манипулированию
2. Тенденция к манипулированию
А) Первые систематические исследования тенденции к манипулированию были осуществлены Харлоу1. Обе его работы были проведены на макаках-резусах, а в качестве предметов для манипулирования использовались механические головоломки, состоявшие либо из 3, либо из 6 элементов (рис. 1). Первый эксперимент (с головоломкой из 3 элементов) показал, что в течение 14 опытных дней обезьяна с интересом манипулировала головоломкой и освоила все операции, с помощью которых головоломка разбирается, не получая за это никакого вознаграждения извне. Во втором эксперименте Харлоу подверг еще более детальному изучению процесс и возможности научения и динамику интереса обезьяны к этому виду манипуляций.
Эксперимент проходил следующим образом. Механическая головоломка (рис. 1) находилась в клетке обезьяны, расположенной в свою оче-
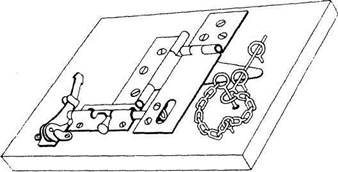
Рис. 1. Механическая головоломка из 6 элементов.
Головоломка, применявшаяся в первом исследовании2,
была проще и состояла из 3 элементов3
редь в закрытом помещении. Обезьяна все время (12 дней) имела доступ во внешнюю и значительно большую клетку, соединенную с первой. Пять раз в день (почти каждые 2 ч) экспериментатор входил в клетку, чтобы
 1 См.: Harlow H.F.,Harlow M.K.,Meyer D.R. Learning motivated by a manipulation
1 См.: Harlow H.F.,Harlow M.K.,Meyer D.R. Learning motivated by a manipulation
drive // J. Exp. Psychol. 1950. 40. P. 228-234; Harlow H.F. Learning and satiation of
response in intrinsically motivated complex puzzle performance by monkeys // J. Сотр.
Physiol. Psychol. 1950. 43. P. 289-294.
2 См.: Harlow H.F..Harlow M.K.,Meyer D.R. Learning motivated by a manipulation
drive // J. Exp. Psychol. 1950. 40. P. 228-234.
3 См.: Harlow H.F. Learning and satiation of response in intrinsically motivated complex
puzzle performance by monkeys // J. Сотр. Physiol. Psychol. 1950. 43. P. 290.
Тема 12. Психология мотивации

|
I o.o-i
0,5-
«1,0-
«a a
§■2,0-
2.5-
| 3,0 |
о
6 8 (0 12
Дна
Рис. 2. Кривая научения (отношение неправильных ответов к правильным) для 5 элементов механической головоломки за 12 дней эксперимента Головоломка состояла фактически из 5 элементов, шестой — шарнирная петля — был несъемным. Результаты 1-го, 2-го и 4-го дней значимо отличаются от результатов 10-го, 11-го и 12-го дней, р = 0,01. В конце каждого 2-часового сеанса экспериментатор входил в клетку и фиксировал число разобранных к этому моменту механизмов. Величина активности обезьян в ненаблюдаемый период определялась путем вычитания из этого числа количества механических элементов, разобранных в течение первых 5 мин. Эта величина также возрастала в течение 12 дней1
снова собрать головоломку, которую обезьяна успела разобрать в перерыве между его посещениями. В течение первых 5 мин каждого периода экспериментатор наблюдал за поведением обезьяны. Он отмечал, в частности, число правильных и неправильных манипуляций с игрушкой, проделанных обезьяной за это время. Действия считались правильными, если они приводили к разбору или освобождению одного из элементов головоломки, и неправильными, если обезьяна манипулировала с элементами не в установленном порядке или не могла их освободить.
На рис. 2 представлена кривая успехов двух обезьян, принимавших участие в опытах в течение 12 дней подряд; по ординате откладывается отношение неправильных действий к правильным. Кривая показывает, что научение обезьян происходит, несмотря на отсутствие какого-либо вознаграждения (пищи, например) извне. Другие графики автора свидетельствуют о том, что эти две обезьяны осуществляли соответственно 63 и 82 правильных действия в течение первых шести дней и 157 и 134 в течение последующих шести дней (за первые пять минут каждого сеанса). Интенсивность мотивации и активности обезьяны проявлялась, в частности, в том, что в последние 6 дней 81% головоломок разбирались практически в первые 5 мин после предъявления их животному.
На 13-й день Харлоу изучал феномен насыщения — каждые 6 мин в течение 10 ч (всего 100 сеансов) он предлагал каждой из обезьян вновь собранную головоломку. Он отметил заметное сокращение числа манипуляций, что свидетельствует об эффекте насыщения. Однако каждая из двух обезьян разобрала почти половину из возможного числа механизмов
 1 См.: Harlow H.F. Learning and satiation of response in intrinsically motivated complex puzzle performance by monkeys // J. Сотр. Physiol. Psychol. 1950. 43. P. 292.
1 См.: Harlow H.F. Learning and satiation of response in intrinsically motivated complex puzzle performance by monkeys // J. Сотр. Physiol. Psychol. 1950. 43. P. 292.
Нюттен Ж. [Познавательная и социальная мотивации] 117
 (100X6), а именно 286, что говорит об удивительной активности обезьяны и предполагает сохранение связанных с ней мотивов.
(100X6), а именно 286, что говорит об удивительной активности обезьяны и предполагает сохранение связанных с ней мотивов.
Б) Первичная потребность или производная тенденция? Пытаясь найти объяснение этой очень сильной мотивации, авторы предположили в своей первой работе1, что она имеет социальный характер из-за присутствия экспериментатора в начале каждого сеанса. Однако в конечном счете они пришли к отрицательному выводу и даже обнаружили, что у двух из четырех участвовавших в эксперименте обезьян уход экспериментатора в большей степени стимулировал активность животных, чем его присутствие. Возможность вторичного подкрепления, связанного с потребностью в пище, также исключается. Таким образом, авторам пришлось признать существование особой потребности в манипулировании. Харлоу2 обнаружил также, что удаление некоторых участков головного мозга приводит к снижению активности, связанной с манипулированием. Некоторые исследователи из школы Халла тем не менее подвергают сомнению специфический и первичный характер потребности в исследовании и манипулировании3. Следует отметить также, что до сих пор ни один из ученых не изучил происхождения и развития этой потребности у животных. С другой стороны, до сих пор точно не известны те свойства, которые придают новому стимулу положительную валентность для человека и животного. О более поздних исследованиях в этой области сообщается в работах Биндры и Спиннера, Карра и Брауна, Эрлиха4.
В) Амбивалентность необычного и неизвестного. При некоторых условиях неизвестное, новое или необычное вызывает негативную реакцию, причем ярко выраженные реакции страха наблюдаются значительно чаще, чем реакции приближения. В отдельных случаях имеет место любопытное сочетание притягательности и страха — тот соблазн опасного, но увлекательного приключения, который столь характерен для некоторых детских игр и поступков взрослых, стремящихся исследовать неизвестное. Виз5 изучал негативные реакции ребенка при встрече с чужими или странными людьми. Он проследил развитие таких реакций от полного двигательного торможения и перевозбуждения до постепенного
 1 См.: Harlow H.F.,Harlow M.К.,Meyer D.R. Learning motivated by a manipulation
1 См.: Harlow H.F.,Harlow M.К.,Meyer D.R. Learning motivated by a manipulation
drive // J. Exp. Psychol. 1950. 40. P. 232-233.
2 См.: Harlow H.F. Learning and satiation of response in intrinsically motivated complex
puzzle performance by monkeys // J. Сотр. Physiol. Psychol. 1950. 43. P. 289-294.
3 См., например: Brown J.S. The motivation of behavior. N. Y.: McGraw-Hill Book,
1961. P. 331-339.
4 См.: Bindra D., Spinner N. Response to different degrees of novelty: the incidence of
various activities // J. Exp. Anal. Behav. 1958. 1. P. 341-350; Carr R.M., Brown W.L. The
effect of sustained novelty upon manipulation on rhesus monkeys // J. Genet. Psychol.
1959. 61. P. 121-125; Ekrlich A. Effect of past experience on exploratory behavior in rats
// Canad. J. Psychol. 1959. 13. P. 248-254.
5 См.: Lewin K. A dynamic theory of personality. Selected papers. N. Y: McGraw-Hill,
1935. P. 261-264.
118 Тема 12. Психология мотивации
 привыкания в зависимости от расстояния до незнакомого человека и времени его присутствия и от того, находится ли ребенок в поле зрения или позади этого человека. С другой стороны, Хебб и Ризен1 изучали реакции страха у обезьяны, оказавшейся в незнакомой или новой обстановке. Выросший в изоляции шимпанзе не испытывает страха при первой встрече с человеком, хотя человеческое лицо, несомненно, является для животного «необычным» объектом. Но у той же самой обезьяны обнаруживаются реакции страха при виде нового человека после того, как она познакомилась с рядом людей и научилась их узнавать. Это нельзя объяснить ни образованием условных связей, ни генерализацией опасного стимула, поскольку до сих пор человек был для молодой обезьяны источником лишь приятных ощущений. Страх возникал у шимпанзе даже в том случае, когда кто-либо из известных уже ей людей приближался к клетке в одежде другого, также знакомого ей человека. На основании этих фактов Хебб2 сформулировал следующую гипотезу: расхождение между тем, что воспринимают рецепторы в данный момент, и следами того, что было воспринято ранее, приводит к определенному нарушению мозговых процессов, в результате возникает эмоция страха. Некоторые эксперименты3 поставили под сомнение универсальность этой гипотезы. Все это говорит о том, что проблема валентности неизвестного и непривычного по-прежнему остается нерешенной.
привыкания в зависимости от расстояния до незнакомого человека и времени его присутствия и от того, находится ли ребенок в поле зрения или позади этого человека. С другой стороны, Хебб и Ризен1 изучали реакции страха у обезьяны, оказавшейся в незнакомой или новой обстановке. Выросший в изоляции шимпанзе не испытывает страха при первой встрече с человеком, хотя человеческое лицо, несомненно, является для животного «необычным» объектом. Но у той же самой обезьяны обнаруживаются реакции страха при виде нового человека после того, как она познакомилась с рядом людей и научилась их узнавать. Это нельзя объяснить ни образованием условных связей, ни генерализацией опасного стимула, поскольку до сих пор человек был для молодой обезьяны источником лишь приятных ощущений. Страх возникал у шимпанзе даже в том случае, когда кто-либо из известных уже ей людей приближался к клетке в одежде другого, также знакомого ей человека. На основании этих фактов Хебб2 сформулировал следующую гипотезу: расхождение между тем, что воспринимают рецепторы в данный момент, и следами того, что было воспринято ранее, приводит к определенному нарушению мозговых процессов, в результате возникает эмоция страха. Некоторые эксперименты3 поставили под сомнение универсальность этой гипотезы. Все это говорит о том, что проблема валентности неизвестного и непривычного по-прежнему остается нерешенной.
|
|
|
