 |


Гены-супрессоры клеточного деления (антионкогены).
|
|
|
|
Гены-супрессоры клеточного деления (антионкогены).
· Они действуют как ингибиторы проведения ростостимулирующих сигналов в клетке, кодируют белки, блокирующие пролиферацию клеток, предупреждая тем самым возможность нерегулируемой пролиферации.
· Инактивация антионкогенов, вызванная их мутациями (точечными или делециями), приводит к неконтролируемому росту клеток
· Для выключения гена-супрессора необходимы две мутации в обоих его аллелях. Наличие первой мутации в одном из аллелей антионкогена предрасполагает к возникновению опухоли, и если такой мутантный аллель унаследован, то достаточно второй соматической мутации, чтобы произошла опухолевая трансформация клетки.
Гены-супрессоры осуществляют роль негативных регуляторов прохождения клетки по клеточному циклу, конечным результатом которого является митоз.
Клеточный цикл разделяется на 4 дискретных временных периода: G1, S, G2 и М.
· В фазе S (синтетическая фаза) происходит репликация ДНК, фаза М - митоз.
· Временной механизм прохождения клеткой этого цикла контролируется синтезом и распадом специальных белков - циклинов.
· Их экспрессия периодически возрастает в течение одной фазы клеточного цикла и затем снижается в другой фазе.
· Циклины группы В накапливаются в G2-фазе и распадаются в М, циклины Е и D действуют в G1-фазе, циклины А - в S-фазе.
· Циклины образуют комплексы с так называемыми циклинзависимыми протеин-киназами (ЦЗПК) разных типов. Образование комплексов циклина Е с ЦЗПК-2 и циклина D с ЦЗПК-4 вызывает фосфорилирование нескольких белков, необходимых для вхождения клетки в S-фазу цикла.
· Одним из таких белков является продукт гена-супрессора Rb, с инактивацией которого связано развитие ретинобластомы в раннем детском возрасте и наследственной формы остеосаркомы. Нефосфорилированная форма белка антионкогена Rb связывает транскрипционные факторы генов, регулирующих репликацию ДНК в S-фазе, что препятствует дальнейшему продвижению находящихся в G1-фазе клеток по циклу.
|
|
|
· При фосфорилировании белка Rb транскрипционные факторы освобождаются, активируются и вызывают вступление клетки в S-фазу.
· Таким образом, в норме антионкоген Rb осуществляет негативный контроль пролиферации, разрешая или не разрешая клеткам вхождение в фазу репликации ДНК (S-фазу клеточного цикла). Утрата этой функции в результате инактивации антионкогена Rb (его мутации) приводит к тому, что транскрипционные факторы остаются несвязанными и клетка безостановочно «пробегает» по циклу даже в тех случаях, когда действует запрет на пролиферацию.
Другим антионкогеном, выполняющим в клетке росторегулирующую функцию, является p53.
· Белок, кодируемый этим антионкогеном, локализован в ядре и является транскрипционным фактором, который включает ряд генов, в том числе и ген waf1.
· Продукт последнего инактивирует комплексы циклина Е с ЦЗПК-2 и циклина D с ЦЗПК-4, необходимые для вхождения клетки в S-фазу цикла, в результате чего клетка задерживается в G1-фазе.
· Мутация р53, как и Rb, приводит к нарушению этой регуляции и к безостановочному делению клетки. В норме р53 препятствует вхождению в S-фазу клеток с поврежденной или измененной ДНК, поддерживая таким образом целостность клеточного генома.
Эту функцию он реализует двумя способами:
1) временно задерживая клетку в G1-фазе цикла, давая ей возможность исправить повреждения ДНК прежде, чем клетка вступит в S-фазу;
2) запуская механизм апоптоза (программируемой гибели клетки) в тех случаях, когда повреждения ДНК столь серьезны, что не подлежат исправлению.
|
|
|
Индукция апоптоза в аномальных клетках - чрезвычайно важная функция антионкогена р53. Благодаря апоптозу исключается возможность передачи серьезных поломов ДНК в поколениях клеток. В случае нарушения этой функции р53 происходит накопление клеток с различными хромосомными повреждениями, что является характерным для клеток опухолей.
Мутации р53 выявляются примерно в 60% злокачественных опухолей у человека. У людей с врожденной мутацией одного из аллелей р53 опухоли (саркомы, лимфолейкозы, рак молочной железы) обнаруживаются в молодом возрасте с вероятностью 100% (семейный синдром Ли-Фраумени). Именно р53 во многом определяет реакции опухоли на химио-и (или) лучевую терапию. В тех опухолях, где р53 не изменен и нормально функционирует, повреждение ДНК под влиянием химиотерапии или облучения вызывает апоптоз опухолевых клеток. В опухолях с инактивированным р53 индуцированные повреждения ДНК не приводят к апоптозу, и такие опухоли резистентны к химио- и лучевой терапии.
Из других генов-супрессоров, с инактивацией которых связывают возникновение определенных опухолей, можно назвать АРС (семейный аденоматозный полипоз толстого кишечника), WT1 (опухоль Вильмса), NF1 (нейрофиброматоз Реклингаузена) и некоторые другие, которые представлены в табл. 14. 5.
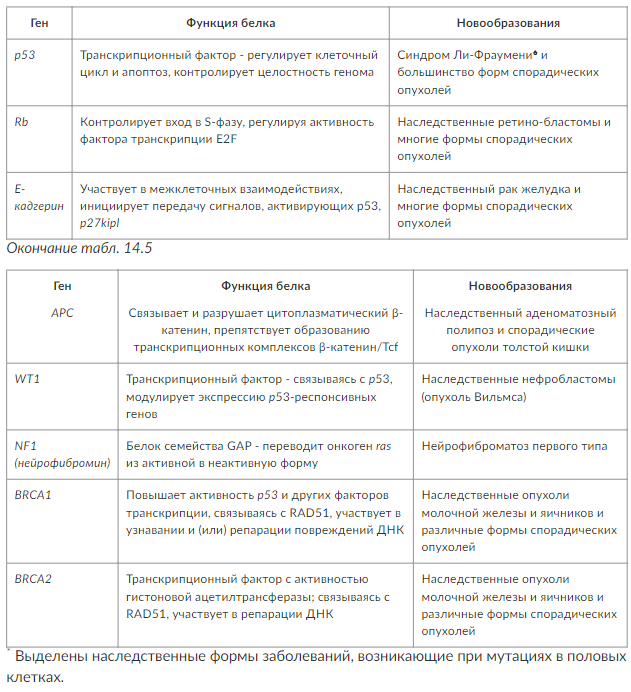
Таким образом, для появления опухоли необходимо, чтобы в одной и той же клетке мутации формировались в нескольких разных генах (протоонкогенах с превращением их в онкогены, генах-супрессорах и др. ). Опухоли одного и того же клеточного или тканевого типа могут развиваться в результате различных комбинаций мутаций в различных генах. Следствием этих генетических изменений является утрата контроля над клеточной пролиферацией. В итоге клетка приобретает трансформированный фенотип, включающий в себя не только нерегулируемую пролиферацию, но и целый ряд характерных изменений структуры и обмена веществ. Например, считается, что развитие злокачественной опухоли толстого кишечника требует от 7 до 10 генетических событий в 4-5 различных генах.
|
|
|
