 |


Смирнов В.М., Будылина С.М. Физиология сенсорных систем и высшая нервная деятельность: Учеб. пособие для студ. высш. учеб. заведений. – М.: Академия, 2003. – с. 35 – 67
|
|
|
|
Зрительный анализатор
Зрительный анализатор представляет собой совокупность структур, воспринимающих световую энергию в виде электромагнитного излучения с длиной волны 400 – 700 нм и дискретных частиц фотонов, или квантов, и формирующих зрительные ощущения. С помощью глаза воспринимается 80 – 90 % всей информации об окружающем мире.

Рисунок: Орган зрения
Благодаря деятельности зрительного анализатора различают освещенность предметов, их цвет, форму, величину, направление передвижения, расстояние, на которое они удалены от глаза и друг от друга. Все это позволяет оценивать пространство, ориентироваться в окружающем мире, выполнять различные виды целенаправленной деятельности.
Наряду с понятием зрительного анализатора существует понятие органа зрения.
Орган зрения — это глаз, включающий три различных в функциональном отношении элемента:
1) глазное яблоко, в котором расположены световоспринимающий, светопреломляющий и светорегулирующий аппараты;
2) защитные приспособления, т. е. наружные оболочки глаза (склера и роговица), слезный аппарат, веки, ресницы, брови;
3) двигательный аппарат, представленный тремя парами глазных мышц (наружная и внутренняя прямые, верхняя и нижняя прямые, верхняя и нижняя косые), которые иннервируются III (глазодвигательный нерв), IV (блоковый нерв) и VI (отводящий нерв) парами черепных нервов.
Структурно-функциональная характеристика
Рецепторный (периферический) отдел зрительного анализатора (фоторецепторы) подразделяется на палочковые и колбочковые нейросенсорные клетки, наружные сегменты которых имеют соответственно палочковидную («палочки») и колбочковидную («колбочки») формы. У человека насчитывается 6 – 7 млн колбочек и 110 – 125 млн палочек.
|
|
|
Место выхода зрительного нерва из сетчатки не содержит фоторецепторов и называется слепым пятном. Латерально от слепого пятна в области центральной ямки лежит участок наилучшего видения — желтое пятно, содержащее преимущественно колбочки. К периферии сетчатки число колбочек уменьшается, а число палочек возрастает, и периферия сетчатки содержит одни лишь палочки.
Различия функций колбочек и палочек лежит в основе феномена двойственности зрения. Палочки являются рецепторами, воспринимающими световые лучи в условиях слабой освещенности, т.е. бесцветное, или ахроматическое, зрение. Колбочки же функционируют в условиях яркой освещенности и характеризуются разной чувствительностью к спектральным свойствам света (цветное или хроматическое зрение). Фоторецепторы обладают очень высокой чувствительностью, что обусловлено особенностью строения рецепторов и физико-химических процессов, лежащих в основе восприятия энергии светового стимула. Полагают, что фоторецепторы возбуждаются при действии на них 1 – 2 квантов света.
Палочки и колбочки состоят из двух сегментов – наружного и внутреннего, которые соединяются между собой посредством узкой реснички. Палочки и колбочки ориентированы в сетчатке радиально, а молекулы светочувствительных белков расположены в наружных сегментах таким образом, что около 90 % их светочувствительных групп лежат в плоскости дисков, входящих в состав наружных сегментов. Свет оказывает наибольшее возбуждающее действие в том случае, если направление луча совпадает с длинной осью палочки или колбочки, при этом он направлен перпендикулярно дискам их наружных сегментов.
Фотохимические процессы в сетчатке глаза. В рецепторных клетках сетчатки находятся светочувствительные пигменты (сложные белковые вещества) – хромопротеиды, которые обесцвечиваются на свету. В палочках на мембране наружных сегментов содержится родопсин, в колбочках – йодопсин и другие пигменты.
|
|
|
Родопсин и йодопсин состоят из ретиналя (альдегида витамина А1) и гликопротеида (опсина). Имея сходство в фотохимических процессах, они различаются тем, что максимум поглощения находится в различных областях спектра. Палочки, содержащие родопсин, имеют максимум поглощения в области 500 нм. Среди колбочек различают три типа, которые отличаются максимумами в спектрах поглощения: одни имеют максимум в синей части спектра (430 – 470 нм), другие в зеленой (500 – 530), третьи – в красной (620 – 760 нм) части, что обусловлено наличием трех типов зрительных пигментов. Красный кол бочковый пигмент получил название «йодопсин». Ретиналь может находиться в различных пространственных конфигурациях (изомерных формах), но только одна из них – 11-ЦИС-изомер ретиналя выступает в качестве хромофорной группы всех известных зрительных пигментов. Источником ретиналя в организме служат каротиноиды.
Фотохимические процессы в сетчатке протекают весьма экономно. Даже при действии яркого света расщепляется только небольшая часть имеющегося в палочках родопсина (около 0,006 %).
В темноте происходит ресинтез пигментов, протекающий с поглощением энергии. Восстановление йодопсина протекает в 530 раз быстрее, чем родопсина. Если в организме снижается содержание витамина А, то процессы ресинтеза родопсина ослабевают, что приводит к нарушению сумеречного зрения, так называемой куриной слепоте. При постоянном и равномерном освещении устанавливается равновесие между скоростью распада и ресинтеза пигментов. Когда количество света, падающего на сетчатку, уменьшается, это динамическое равновесие нарушается и сдвигается в сторону более высоких концентраций пигмента. Этот фотохимический феномен лежит в основе темновой адаптации.
Особое значение в фотохимических процессах имеет пигментный слой сетчатки, который образован эпителием, содержащим фусцин. Этот пигмент поглощает свет, препятствуя отражению и рассеиванию его, что обусловливает четкость зрительного восприятия. Отростки пигментных клеток окружают светочувствительные членики палочек и колбочек, принимая участие в обмене веществ фоторецепторов и в синтезе зрительных пигментов.
|
|
|
Вследствие фотохимических процессов в фоторецепторах глаза при действии света возникает рецепторный потенциал, который представляет собой гиперполяризацию мембраны рецептора. Это отличительная черта зрительных рецепторов, активация других рецепторов выражается в виде деполяризации их мембраны. Амплитуда зрительного рецепторного потенциала увеличивается при увеличении интенсивности светового стимула. Так, при действии красного цвета, длина волны которого составляет 620 – 760 нм, рецепторный потенциал более выражен в фоторецепторах центральной части сетчатки, а синего (430 – 470 нм) – в периферической.
Синаптические окончания фоторецепторов конвергируют на биполярные нейроны сетчатки. При этом фоторецепторы центральной ямки связаны только с одним биполяром.
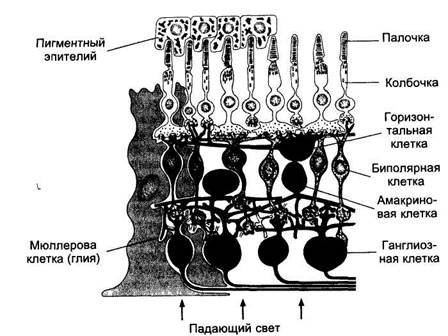
Рисунок: Схема строения сетчатки
Проводниковый отдел. Первый нейрон проводникового отдела зрительного анализатора представлен биполярными клетками сетчатки.
Считают, что в биполярных клетках возникают потенциалы действия подобно рецепторным и горизонтальным НС. В одних биполярах на включение и выключение света возникает медленная длительная деполяризация, а в других — на включение — гиперполяризация, на выключение — деполяризация.
Аксоны биполярных клеток в свою очередь конвергируют на ган-глиозные клетки (второй нейрон). В результате на каждую ганглиозную клетку могут конвергировать около 140 палочек и 6 колбочек, при этом чем ближе к желтому пятну, тем меньше фоторецепторов конвергирует на одну клетку. В области желтого пятна конвергенция почти не осуществляется и количество колбочек почти равно количеству биполярных и ганглиозных клеток. Именно это объясняет высокую остроту зрения в центральных отделах сетчатки.
Периферия сетчатки отличается большой чувствительностью к слабому свету. Это обусловлено, по-видимому, тем, что до 600 палочек конвергируют здесь через биполярные клетки на одну и ту же ганглиозную клетку. В результате сигналы от множества палочек суммируются и вызывают более интенсивную стимуляцию этих клеток.
|
|
|
[…]
Проводниковый отдел, начинающийся в сетчатке (первый нейрон — биполярный, второй нейрон — ганглиозные клетки), анатомически представлен далее зрительными нервами и после частичного перекреста их волокон — зрительными трактами. В каждом зрительном тракте содержатся нервные волокна, идущие от внутренней (носовой) поверхности сетчатки глаза одноименной стороны и от наружной половины сетчатки другого глаза. Волокна зрительного тракта направляются к зрительному бугру (собственно таламур), к метаталамусу (наружные коленчатые тела) и к ядрам подушки. Здесь расположены третий нейрон зрительного анализатора. От них зрительные нервные волокна направляются в кору полушарий большого мозга.
В наружных (или латеральных) коленчатых телах, куда приходят волокна из сетчатки, есть рецептивные поля, которые также имеют округлую форму, но меньше по размеру, чем в сетчатке. Ответы нейронов здесь носят фазический характер, но более выражены, чем в сетчатке. На уровне наружных коленчатых тел происходит процесс взаимодействия афферентных сигналов, идущих от сетчатки глаза, с эфферентными из области коркового отдела зрительного анализатора. С участием ретикулярной формации здесь происходит взаимодействие со слуховой и другими сенсорными системами, что обеспечивает процессы избирательного зрительного внимания путем выделения наиболее существенных компонентов сенсорного сигнала.
Центральный, или корковый отдел зрительного анализатора расположен в затылочной доле (поля 17, 18, 19 по Бродману) или VI, V2, V3 (согласно принятой номенклатуре). Считают, что первичная проекционная область (поле 17) осуществляет специализированную, но более сложную, чем в сетчатке и в наружных коленчатых телах, переработку информации. Рецептивные поля нейронов зрительной коры небольших размеров имеют вытянутые, почти прямоугольные, а не округлые формы. Наряду с этим имеются сложные и сверхсложные рецептивные поля детекторного типа. Эта особенность позволяет выделять из цельного изображения лишь отдельные части линий с различным расположением и ориентацией, при этом проявляется способность избирательно реагировать на эти фрагменты.
В каждом участке коры сконцентрированы нейроны, которые образуют колонку, проходящую по глубине через все слои вертикально, при этом происходит функциональное объединение нейронов, выполняющих сходную функцию. Разные свойства зрительных объектов (цвет, форма, движение) обрабатываются в разных частях зрительной коры большого мозга параллельно.
|
|
|
[…]
Механизмы, обеспечивающие ясное видение в различных условиях
При рассмотрении объектов, находящихся на разном удалении от наблюдателя, ясному видению способствуют следующие процессы.
1. Конвергенционные и дивергенционные движения глаз, благодаря которым осуществляется сведение или разведение зрительных осей. Если оба глаза двигаются в одном направлении, такие движения называются содружественными.
2. Реакция зрачка, которая происходит синхронно с движением глаз. Так, при конвергенции зрительных осей, когда рассматриваются близко расположенные предметы, происходит сужение зрачка, т. е. конвергентная реакция зрачков. Эта реакция способствует уменьшению искажения изображения, вызываемого сферической аберрацией. Сферическая аберрация обусловлена тем, что преломляющие среды глаза имеют неодинаковое фокусное расстояние в разных участках. Центральная часть, через которую проходит оптическая ось, имеет большее фокусное расстояние, чем периферическая часть. Поэтому изображение на сетчатке получается нерезким. Чем меньше диаметр зрачка, тем меньше искажения, вызываемые сферической аберрацией. Конвергентные сужения зрачка включают в действие аппарат аккомодации, обусловливающий увеличение преломляющей силы хрусталика.
Зрачок является также аппаратом устранения хроматической аберрации, которая обусловлена тем, что оптический аппарат глаза, как и простые линзы, преломляет свет с короткой волной сильнее, чем с длинной волной. Исходя из этого, для более точкой фокусировки предмета красного цвета требуется большая степень аккомодации, чем для синего. Именно поэтому синие предметы кажутся более удаленными, чем красные, будучи расположенными на одном и том же расстоянии.
3. Аккомодация является главным механизмом, обеспечивающим ясное видение разноудаленных предметов, и сводится к фокусированию изображения от далеко или близко расположенных предметов на сетчатке. Основной механизм аккомодации заключается в непроизвольном изменении кривизны хрусталика глаза.
Благодаря изменению кривизны хрусталика, особенно передней поверхности, его преломляющая сила может меняться в пределах 10 — 14 диоптрий. Хрусталик заключен в капсулу, которая по краям (вдоль экватора хрусталика) переходит в фиксирующую хрусталик связку (циннова связка), в свою очередь соединенную с волокнами ресничной (цилиарной) мышцы. При сокращении цилиарной мышцы натяжение цинновых связок уменьшается, а хрусталик вследствие своей эластичности становится более выпуклым. Преломляющая сила глаза увеличивается, и глаз настраивается на видение близко расположенных предметов. Когда человек смотрит вдаль, циннова связка находится в натянутом состоянии, что приводит к растягиванию сумки хрусталика и его утолщению. Иннервация цилиарной мышцы осуществляется симпатическими и парасимпатическими нервами. Импульсация, поступающая по парасимпатическим волокнам глазодвигательного нерва, вызывает сокращение мышцы. Симпатические волокна, отходящие от верхнего шейного узла, вызывают ее расслабление. Изменение степени сокращения и расслабления цилиарной мышцы связано с возбуждением сетчатки и находится под влиянием коры головного мозга. Преломляющая сила глаза выражается в диоптриях (Д). Одна диоптрия соответствует преломляющей силе линзы, главное фокусное расстояние которой в воздухе равно 1 м. Если главное фокусное расстояние линзы равно, например, 0,5 или 2 м, то ее преломляющая сила составляет, соответственно, 2Д или 0,5Д. Преломляющая сила глаза без явления аккомодации равна 58 —60 Д и называется рефракцией глаза.
При нормальной рефракции глаза лучи от далеко расположенных предметов после прохождения через светопреломляющую систему глаза собираются в фокусе на сетчатке в центральной ямке. Нормальная рефракция глаза носит название эмметропии, а такой глаз называют эмметропическим. Наряду с нормальной рефракцией наблюдаются ее аномалии.
Миопия (близорукость) — это такой вид нарушения рефракции, при котором лучи от предмета после прохождения через светопреломляющий аппарат фокусируются не на сетчатке, а впереди нее. Это может зависеть от большой преломляющей силы глаза или от большой длины глазного яблока. Близкие предметы близорукий видит без аккомодации, отдаленные предметы видит неясными, расплывчатыми. Для коррекции применяются очки с рассеивающими двояковогнутыми линзами.
Гиперметропия (дальнозоркость) — вид нарушения рефракции, при котором лучи от далеко расположенных предметов в силу слабой преломляющей способности глаза или при малой длине глазного яблока фокусируются за сетчаткой. Даже удаленные предметы дальнозоркий глаз видит с напряжением аккомодации, вследствие чего развивается гипертрофия аккомодационных мышц. Для коррекции применяют двояковыпуклые линзы.
Астигматизм — вид нарушения рефракции, при котором лучи не могут сходиться в одной точке, в фокусе (от греч. stigme — точка), обусловлен различной кривизной роговицы и хрусталика в различных меридианах (плоскостях). При астигматизме предметы кажутся сплющенными или вытянутыми, его коррекцию осуществляют сфероцилиндрическими линзами.
Следует отметить, что к светопреломляющей системе глаза относятся также: роговица, влага передней камеры глаза, хрусталик и стекловидное тело. Однако их преломляющая сила, в отличие от хрусталика, не регулируется и в аккомодации участия не принимает. После прохождения лучей через преломляющую систему глаза на сетчатке получается действительное, уменьшенное и перевернутое изображение. Но в процессе индивидуального развития сопоставление ощущений зрительного анализатора с ощущениями двигательного, кожного, вестибулярного и других анализаторов, как отмечалось выше, приводит к тому, что человек воспринимает внешний мир таким, какой он есть на самом деле.
Бинокулярное зрение (зрение двумя глазами) играет важную роль в восприятии разноудаленных предметов и определении расстояния до них, дает более выраженное ощущение глубины пространства по сравнению с монокулярным зрением, т. е. зрением одним глазом. При рассматривании предмета двумя глазами его изображение может попадать на симметричные (идентичные) точки сетчаток обоих глаз, возбуждения от которых объединяются в корковом конце анализатора в единое целое, давая при этом одно изображение. Если изображение предмета попадает на неидентичные (диспаратные) участки сетчатки, то возникает раздвоение изображения. Процесс зрительного анализа пространства зависит не только от наличия бинокулярного зрения, существенную роль в этом играют условно-рефлекторные взаимодействия, складывающиеся между зрительным и двигательным анализаторами. Определенное значение имеют конвергенционные движения глаз и процесс аккомодации, которые управляются по принципу обратный связей. Восприятие пространства в целом связано с определением пространственных отношений видимых предметов — их величины, формы, отношения друг к другу, что обеспечивается взаимодействием различных отделов анализатора; значительную роль при этом играет приобретенный опыт.
При движении объектов ясному видению способствуют следующие факторы: 1) произвольные движения глаз вверх, вниз, влево или вправо со скоростью движения объекта, что осуществляется благодаря содружественной деятельности глазодвигательных мышц; 2) при появлении объекта в новом участке поля зрения срабатывает фиксационный рефлекс — быстрое непроизвольное движение глаз, обеспечивающее совмещение изображения предмета на сетчатке с центральной ямкой. При слежении за движущимся объектом происходит медленное движение глаз — следящее движение.
При рассматривании неподвижного предмета для обеспечения ясного видения глаз совершает три типа мелких непроизвольных Движений: тремор — дрожание глаза с небольшой амплитудой и частотой, дрейф — медленное смещение глаза на довольно значительное расстояние и скачки (флики) — быстрые движения глаз. Также существуют саккадические движения (саккады) — содружественные движения обоих глаз, совершаемые с большой скоростью. Наблюдаются саккады при чтении, просматривании картин, когда обследуемые точки зрительного пространства находятся на одном удалении 6т наблюдателя и других объектов. Если заблокировать эти движения глаз, то окружающий нас мир вследствие адаптации рецепторов сетчатки станет трудно различимым, каким он является у лягушки. Глаза лягушки неподвижны, поэтому она хорошо различает только движущиеся предметы, например бабочек. Именно поэтому лягушка приближается к змее, которая постоянно выбрасывает наружу свой язык. Находящуюся в состоянии неподвижности змею лягушка не различает, а ее движущийся язык принимает за летающую бабочку.
В условиях изменения освещенности ясное видение обеспечивают зрачковый рефлекс, темновая и световая адаптация.
Зрачок регулирует интенсивность светового потока, действующего на сетчатку, путем изменения своего диаметра. Ширина зрачка может колебаться от 1,5 до 8,0 мм. Сужение зрачка (миоз) происходит при увеличении освещенности, а также при рассматривании близко расположенного предмета и во сне. Расширение зрачка (мидриаз) происходит при уменьшении освещенности, а также при возбуждении рецепторов, любых афферентных нервов, при эмоциональных реакциях напряжения, связанных с повышением тонуса симпатического отдела нервной системы (боль, гнев, страх, радость и т.д.), при психических возбуждениях (психозы, истерии и т.д.), при удушье, наркозе. Зрачковый рефлекс при изменении освещенности хотя и улучшает зрительное восприятие (в темноте расширяется, что увеличивает световой поток, падающий на сетчатку, на свету сужается), однако главным механизмом все же является темновая и световая адаптация.
Темновая адаптация выражается в повышении чувствительности зрительного анализатора (сенситизация), световая адаптация — в снижении чувствительности глаза к свету. Основу механизмов световой и темновой адаптации составляют протекающие в колбочках и палочках фотохимические процессы, которые обеспечивают расщепление (на свету) и ресинтез (в темноте) фоточувствительных пигментов, а также процессы функциональной мобильности: включение и выключение из деятельности рецепторных элементов сетчатки. Кроме того, адаптацию определяют некоторые нейронные механизмы и, прежде всего, процессы, происходящие в нервных элементах сетчатки, в частности способы подключения фоторецепторов к ганглиозным клеткам с участием горизонтальных и биполярных клеток. Так, в темноте возрастает число рецепторов, подключенных к одной биполярной клетке, и большее их число конвергирует на ганглиозную клетку. При этом расширяется рецептивное поле каждой биполярной и, естественно, ганглиозной клеток, что улучшает зрительное восприятие. Включение же горизонтальных клеток регулируется ЦНС.
Снижение тонуса симпатической нервной системы (десимпатизация глаза) уменьшает скорость темновой адаптации, а введение адреналина оказывает противоположный эффект. Раздражение ретикулярной формации ствола мозга повышает частоту импульсов в волокнах зрительных нервов. Влияние ЦНС на адаптивные процессы в сетчатке подтверждается также тем, что чувствительность неосвещенного глаза к свету изменяется при освещении другого глаза и при действии звуковых, обонятельных или вкусовых раздражителей.
Цветовая адаптация. Наиболее быстрая и резкая адаптация (снижение чувствительности) происходит при действии сине-фиолетового раздражителя. Красный раздражитель занимает среднее положение.
Зрительное восприятие крупных объектов и их деталей обеспечивается за счет центрального и периферического зрения — изменений угла зрения. Наиболее тонкая оценка мелких деталей предмета обеспечивается в том случае, если изображение падает на желтое пятно, которое локализуется в центральной Ямке сетчатки глаза, так как в этом случае имеет место наибольшая острота зрения. Это объясняется тем, что в области желтого пятна располагаются только колбочки, их размеры наименьшие, и каждая колбочка контактирует с малым числом нейронов, что повышает
остроту зрения. Острота зрения определяется наименьшим углом зрения, под которым глаз еще способен видеть отдельно две точки. Нормальный глаз способен различать две светящиеся точки под углом зрения в 1’. Острота зрения такого глаза принимается за единицу. Острота зрения зависит от оптических свойств глаза, структурных особенностей сетчатки и работы нейрональных механизмов проводникового и центрального отделов зрительного анализатора. Определение остроты зрения осуществляется с помощью буквенных или различного вида фигурных стандартных таблиц. Крупные объекты в целом и окружающее пространство воспринимаются в основном за счет периферического зрения, обеспечивающего большое поле зрения.
Поле зрения — пространство, которое можно видеть фиксированным глазом. Различают отдельно поле зрения левого и правого глаз, а также общее поле зрения для двух глаз. Величина поля зрения у людей зависит от глубины положения глазного яблока и формы надбровных дуг и носа. Ахроматическое поле зрения больше хроматического в силу того, что на периферии сетчатки нет рецепторов, воспринимающих цвет (колбочек). В свою очередь, цветное поле зрения неодинаково для различных цветов. Самое узкое поле зрения для зеленого, желтого, больше для красного, еще больше для синего цветов. Величина поля зрения изменяется в зависимости от освещенности. Ахроматическое поле зрения в сумерках увеличивается, на свету уменьшается. Хроматическое поле зрения, наоборот, на свету увеличивается, в сумерках уменьшается. Это зависит от процессов мобилизации и демобилизации фоторецепторов (функциональной мобильности). При сумеречном зрении увеличение количества функционирующих палочек, т.е. их мобилизация, приводит к увеличению ахроматического поля зрения, в то же самое время уменьшение количества функционирующих колбочек (их демобилизация) ведет к уменьшению хроматического поля зрения.
Зрительный анализатор имеет также механизм для различения длины световой волны — цветовое зрение.
Цветовое зрение, зрительные контрасты и последовательные образы
Цветовое зрение — способность зрительного анализатора реагировать на изменения длины световой волны с формированием ощущения цвета. Определенной длине волны электромагнитного излучения соответствует ощущение определенного цвета. Так, ощущение красного цвета соответствует действию света с длиной волны в 620 — 760 нм, а фиолетового — 390 — 450 нм, остальные цвета спектра имеют промежуточные параметры. Смешение всех цветов дает ощущение белого цвета. В результате смешения трех основных цветов спектра — красного, зеленого, сине-фиолетового — в разном соотношении можно получить также восприятие любых других цветов. Ощущение цветов связано с освещенностью. По мере ее уменьшения сначала перестают различаться красные цвета, позднее всех — синие. Восприятие цвета обусловлено в основном процессами, происходящими в фоторецепторах. Наибольшим признанием пользуется трехкомпонентная теория цветоощущения Ломоносова— Юнга—Гельмгольца—Лазарева, согласно которой в сетчатке глаза имеются три вида фоторецепторов — колбочек, раздельно воспринимающих красный, зеленый и сине-фиолетовые цвета. Комбинации возбуждения различных колбочек приводят к ощущению различных цветов и оттенков. Равномерное возбуждение трех видов колбочек дает ощущение белого цвета. Три типа цветочувствительных колбочек были названы модуляторами, колбочки, которые возбуждались при изменении яркости света (четвертый тип), были названы доминаторами. Впоследствии методом микроспектрофотометрии удалось установить, что даже одиночная колбочка может поглощать лучи различной длины волны. Обусловлено это наличием в каждой колбочке различных пигментов, чувствительных к волнам света различной длины.
Несмотря на убедительные аргументы трехкомпонентной теории в физиологии цветового зрения описаны факты, которые не находят объяснения с этих позиций. Это дало возможность выдвинуть теорию противоположных, или контрастных, цветов, т. е. создать так называемую оппонентную теорию цветного зрения Эвальда Геринга.
Согласно этой теории, в глазу и/или в мозге существуют три оппонентных процесса: один — для ощущения красного и зеленого, второй — для ощущения желтого и синего, третий — качественно отличный от двух первых процессов — для черного и белого. Эта теория применима для объяснения передачи информации о цвете в последующих отделах зрительной системы: ганглиозных клетках сетчатки, наружных коленчатых телах, корковых центрах зрения, где функционируют цветооппонентные РП с их центром и периферией.
Таким образом, на основании полученных данных можно полагать, что процессы в колбочках более соответствуют трехкомпонентной теории цветоощущения, тогда как для нейронных сетей сетчатки и вышележащих зрительных центров подходит теория контрастных цветов Геринга.
В восприятии цвета определенную роль играют и процессы, Протекающие в нейронах разных уровней зрительного анализатора (включая сетчатку), которые получили название цветооппонентных нейронов. При действии на глаз излучений одной части спектра они возбуждаются, а другой — тормозятся. Такие нейроны участвуют в кодировании информации о цвете.
Наблюдаются аномалии цветового зрения, которые могут проявляться в виде частичной или полной цветовой слепоты. Людей, вообще не различающих цвета, называют ахроматами. Частичная цветовая слепота имеет место у 8 – 10% мужчин и 0,5% женщин. Полагают, что цветослепота связана с отсутствием у мужчин определенных генов в половой непарной Х-хромосоме. Различаются три вида частичной цветослепоты: протанопия (дальтонизм) — слепота в основном на красный цвет. Этот вид цветослепоты впервые был описан в 1794 году физиком Дж. Дальтоном, у которого наблюдался этот вид аномалии. Людей с таким видом аномалии называют «краснослепыми»; дейтеранопия — понижение восприятия зеленого цвета. Таких людей называют «зеленослепыми»; тританопия — редко встречающаяся аномалия. При этом люди не воспринимают синий и фиолетовый цвета, их называют «фиолетовослепыми».
С точки зрения трехкомпонентной теории цветового зрения каждый из видов аномалии является результатом отсутствия одного из трех колбочковых цветовоспринимающих субстратов. Для диагностики расстройства цветоощущения пользуются цветными таблицами Е.Б. Рабкина, а также специальными приборами, получившими название аномалоскопов. Выявление различных аномалий цветового зрения имеет большое значение при определении профессиональной пригодности человека для различных видов работ (шофера, летчика, художника и др.).
Возможность оценки длины световой волны, проявляющаяся в способности к цветоощущению, играет существенную роль в жизни человека, оказывая влияние на эмоциональную сферу и деятельность различных систем организма. Красный цвет вызывает ощущение тепла, действует возбуждающе на психику, усиливает эмоции, но быстро утомляет, приводит к напряжению мышц, повышению артериального давления, учащению дыхания. Оранжевый цвет вызывает чувство веселья и благополучия, способствует пищеварению. Желтый цвет создает хорошее, приподнятое настроение, стимулирует зрение и нервную систему. Это самый «веселый» цвет. Зеленый цвет действует освежающе и успокаивающе, полезен при бессоннице, переутомлении, понижает артериальное давление, общий тонус организма и является самым благоприятным для человека. Голубой цвет вызывает ощущение прохлады и действует на нервную систему успокаивающе, причем сильнее зеленого (особенно благоприятен голубой цвет для людей с повышенной нервной возбудимостью), больше, чем при зеленом цвете, понижает артериальное давление и тонус мышц. Фиолетовый цвет не столько успокаивает, сколько расслабляет психику. Создается впечатление, что человеческая психика, следуя вдоль спектра от красного к фиолетовому, проходит всю гамму эмоций. На этом основано использование теста Люшера для определения эмоционального состояния организма.
Зрительные контрасты и последовательные образы. Зрительные ощущения могут продолжаться и после того, как прекратилось раздражение. Такое явление получило название последовательных образов. Зрительные контрасты — это измененное восприятие раздражителя в зависимости от окружающего светового или цветового фона. Существуют понятий светового и цветового зрительных контрастов. Явление контраста может проявляться в преувеличении действительной разницы между двумя одновременными или последовательными ощущениями, поэтому различают одновременные и последовательные контрасты. Серая полоска на белом фоне кажется темнее такой же полоски, расположенной на темном фоне. Это пример одновременного светового контраста. Если рассматривать серый цвет на красном фоне, то он кажется зеленоватым, а если рассматривать серый цвет на синем фоне, то он приобретает желтый оттенок. Это явление одновременного цветового контраста. Последовательный цветовой контраст заключается в изменении цветового ощущения при переводе взгляда на белый фон. Так, если долго смотреть на окрашенную в красный цвет поверхность, а затем перевести взор на белую, то она приобретает зеленоватый оттенок. Причиной зрительного контраста являются процессы, которые осуществляются в фоторецепторном и нейрональном аппаратах сетчатки. Основу составляет взаимное торможение клеток, относящихся к разным рецептивным полям сетчатки и их проекциям в корковом отделе анализаторов.
|
|
|
