 |


Парадигма обращается вспять
|
|
|
|
Первым обрушился камень, на котором держались представления о «первично амитохондриальном» статусе Archezoa. Если помните, предполагалось, что архезои никогда не имели митохондрий. Но чем больше генов разных представителей Archezoa секвенировали исследователи, тем вероятнее казалось, что предполагаемые предки эукариотических клеток, такие как возбудитель амебной дизентерии Entamoeba histolytica, все-таки не были самыми древними представителями этой группы. Нашлись более древние представители — но как раз у них митохондрии были. К сожалению, методы генетической датировки довольно приблизительны и результаты, получаемые с их помощью, бывают ошибочны, а значит, допускают разные толкования. Однако если проведенные датировки были верны, то их результат мог значить только одно: предки Entamoeba histolytica имели митохондрии, а энтамебы утратили их. Если Archezoa — это группа примитивных эукариот, никогда не имевших митохондрий, то E. histolytica — не архезой.
В 1995 г. Грэм Кларк из Национального института здравоохранения (США) и Эндрю Роджер из Университета Далхаузи (Канада) решили еще раз присмотреться к E. histolytica и проверить, нет ли каких-нибудь признаков того, что у нее когда-то были митохондрии. Такие признаки нашлись. В глубинах ядерного генома обнаружились два гена, которые, судя по их нуклеотидным последовательностям, скорее всего, имели митохондриальное происхождение. Вероятно, они попали в ядро клетки-хозяина на самых ранних этапах симбиоза с митохондриями, а затем клетка утратила не только митохондрии, но и все следы их прежнего наличия. Надо заметить, что перенос генов из митохондрий в ядро клетки-хозяина — это нормально, а почему это так, мы узнаем в третьей части книги. Современные митохондрии сохранили лишь небольшое количество генов, все остальные либо совсем потерялись, либо оказались в ядре. Белки, которые кодируются такими генами, часто предназначены для митохондрий. Интересно, что У E. histolytica на самом деле есть странные овальные органеллы, которые могут быть искореженными митохондриями; они похожи на митохондрии по размеру и форме, а несколько выделенных из них белков встречаются в митохондриях других организмов.
|
|
|
Неудивительно, что после этого открытия в центре внимания оказались другие первично амитохондриальные группы. Может быть, у них тоже когда-то были митохондрии? Были проведены аналогичные исследования, и оказалось, что все проверенные таким образом «архезои» когда-то имели митохондрии, а потом утратили их. У Giardia к тому же нашлись органеллы (митосомы), которые, по всей видимости, являются рудиментами митохондрии и продолжают выполнять некоторые их функции (хотя и не главную — аэробное дыхание). Самые удивительные результаты были получены на микроспоридиях. Эта якобы древняя амитохондриальная группа не только когда-то имела митохондрии, но и оказалась вовсе не древней. Их ближайшие родственники — высшие грибы — относительно молодая группа эукариот. Кажущаяся древность микроспоридий — артефакт, связанный с внутриклеточным паразитизмом. А то, что они заражают представителей очень разных групп — не более чем свидетельство их эволюционного успеха.
Нельзя исключать, что настоящие архезои все еще ждут своего первооткрывателя, но общее мнение сходится на том, что это фантомная группа. Все эукариоты, исследованные на данный момент, либо имеют митохондрии, либо когда-то имели их. Согласно имеющимся данным нет никаких оснований полагать, что примитивные архезои когда-либо существовали. А если так, то симбиоз с митохондриями восходит к самому началу эволюционной линии эукариот, а возможно, неотделим от загадки их происхождения: этот симбиоз и был тем уникальным событием, которое дало начало эукариотам.
|
|
|
Если первые эукариоты не были архезоями — иными словами, простыми клетками, питавшимися за счет фагоцитоза, — то на что же они были похожи? Возможно, ответ надо искать в подробном анализе последовательностей ДНК современных эукариот. Мы уже видели, что сравнение нуклеотидных последовательностей позволяет выявить бывшие митохондриальные гены; может быть, тем же путем можно выявить и гены, унаследованные от первой клетки-хозяина. Сама идея проста. Мы знаем, что митохондрии родственны определенной группе бактерий, α-протеобактериям, а значит, можем исключить гены, которые, как кажется, происходят из этого источника, а потом посмотреть, откуда взялись все остальные. Часть этих остальных генов явно должна быть характерна только для эукариот, ведь они эволюционировали на протяжении последних двух тысяч миллионов лет после того, как приобрели митохондрии, а еще некоторая часть могла попасть в геном из других источников. Но даже если и так, мы должны найти хотя бы несколько генов первичной клетки-хозяина. Их унаследовали все потомки, и, естественно, в них должны были накопиться изменения, но по крайней мере некоторое сходство с предком должно было остаться.
Такой подход использовали Мария Ривера и ее коллеги из Калифорнийского университета (Лос-Анджелес, США). Статья с результатами их исследований вышла в 1998 г., а в 2004 г. в Nature был напечатан ее развернутый вариант. Эта группа исследователей сравнила полные геномные последовательности представителей всех трех доменов живых организмов и обнаружила, что у эукариот есть два четко отличающихся класса генов, которые они назвали информационными и операционными генами. Информационные гены кодируют весь базовый аппарат, отвечающий за наследственность, позволяя клетке копировать и транскрибировать ДНК, воспроизводить себя и синтезировать белки. Операционные гены кодируют белки, отвечающие за «повседневную жизнь» — производство энергии и основных строительных блоков жизни, таких как липиды и аминокислоты. Интересно, что почти все операционные гены унаследованы от α-протеобактерий (надо полагать, через митохондрии), но единственный настоящий сюрприз заключался в том, что этих генов оказалось намного больше, чем предполагалось раньше, то есть генетический вклад предка митохондрий был больше, чем мы думали. Но самое удивительное — происхождение информационных генов. Как и предполагалось, они похожи на гены архей, но больше всего они похожи на гены совершенно неожиданной группы архей — метаногенов, этих болотных жителей, которые не любят кислород и образуют метан.
|
|
|
Это не единственная улика, наводящая подозрения на метаногены. Джон Рив с коллегами из университета штата Огайо (город Колумбус) показали, что гистоны (белки, в которые упакована ДНК) очень похожи у эукариот и метаногенов. Это сходство не случайно. Очень похожа не только структура самих гистонов, поразительно сходна и трехмерная конформация, получающаяся при упаковке в них ДНК. Обнаружить одинаковое строение гистонов у двух якобы неродственных организмов, таких как метаногены и эукариоты, это все равно что выяснить, что реактивные двигатели самолетов двух конкурирующих компаний имеют одинаковое устройство. Такое, конечно, бывает, но, согласитесь, трудно поверить, что компании независимо разработали совершенно одинаковый двигатель, не имея доступа ни к чертежам конкурента, ни к исходным чертежам. Скорее всего, одна компания купила или украла проектную документацию у другой. Так и с упаковкой ДНК в гистоны: она настолько похожа у метаногенов и эукариот, что, скорее всего, они получили этот признак от общего предка. Оба варианта построены с использованием одних и тех же исходных чертежей.
Все это складывается в очень внушительную доказательную базу. Улики свидетельствуют о том, что и информационные гены, и гистоны мы унаследовали от метаногенов. Стоило почти смириться с тем, что наш почтенный предок был мерзким паразитом, как выясняется, что на его роль претендует еще более чужеродное нам существо, потомки которого ныне обретаются в смрадных болотах и кишечниках животных. Клетка, вступившая в симбиоз, давший начало эукариотам, была метаногеном.
|
|
|
Теперь мы можем представить, каким именно многообещающим монстром была первая эукариотическая клетка. Это был продукт слияния метаногена (который вырабатывал энергию за счет производства метана) и α-протеобактерии, например паразита вроде Rickettsia. Это удивительный парадокс. Найдется мало организмов, которые ненавидят кислород больше, чем метаногены, разве что некоторые обитатели полностью анаэробных местообитаний. Напротив, мало кто зависит от кислорода больше, чем рикеттсии — эти мелкие паразиты, идеально приспособившиеся к внутриклеточному образу жизни за счет максимального упрощения организации. Они избавились от всех ненужных генов, оставив только гены, нужные для размножения, и гены, нужные для аэробного дыхания. Парадокс заключается вот в чем: если эукариотическая клетка произошла в результате симбиоза между ненавидящими кислород метаногенами и обожающими кислород α-протеобактериями, какую выгоду могло сулить метаногену присутствие бактерии? И, если уж на то пошло, какая польза бактерии от жизни внутри метаногена? И еще: если хозяин не умел фагоцитировать — а метаногены совершенно точно не меняют форму тела и не едят другие клетки, — то как, скажите на милость, будущий симбионт попал внутрь?
Возможно, предложенную Сив Андерссон гипотезу «токсичного кислорода» все же не стоит сбрасывать со счетов. Предположим, что поглощающая кислород бактерия защищала метаноген от токсичного для него кислорода, тем самым позволив ему занять новые местообитания. Но этот сценарий связан с одной серьезной проблемой. Между бактерией и примитивным архезоем, который жил за счет сбраживания органических остатков, такие взаимоотношения действительно могли бы существовать. Архезой выиграл бы от перемещения в любое местообитание, где есть такие остатки. Такие клетки-мусорщики — одноклеточные аналоги шакалов, которые рыщут по африканским саваннам в поисках свежего трупа. Однако такие блуждания убили бы метаноген. Он привязан к местообитанию с низким содержанием кислорода, как бегемот к своему болоту. Метаногены переносят кислород, но не могут производить энергию в его присутствии, потому что им нужен водород, а он крайне редко встречается вместе с кислородом. Так что если метаноген покинет свое местообитание, ему придется поститься до тех пор, пока он не вернется обратно. Гниющие органические остатки его не интересуют, ему лучше там, где нет кислорода. Это означает, что между интересами метаногена, не заинтересованного в смене местообитания, и интересами охочего до кислорода паразита, который не может производить энергию в бескислородной среде, имеются серьезные противоречия.
|
|
|
Этот парадокс усугубляется тем, что, как мы видели, взаимоотношения этих двух организмов не могли быть построены на обмене энергией в форме АТФ — бактерии не имеют экспортеров АТФ и никогда не «делятся» энергией друг с другом. Конечно, не исключено, что «местом встречи» был опять же паразитизм: бактерии поглощали органические продукты жизнедеятельности метаногена изнутри. Однако и тут мы сталкиваемся с противоречиями, так как зависящая от кислорода бактерия не могла производить энергию внутри метаногена, не «убедив» его покинуть уютное бескислородное болото. Представляю себе, как α-протеобактерии гонят стадо метаногенов, как скот на бойню, в богатые кислородом места… но так не бывает. Короче говоря, метаногены погибли бы от голода, покинув свое болото; зависящие от кислорода бактерии погибли бы в этом болоте, а промежуточный вариант — местообитания с небольшим количеством кислорода — был бы в равной мере невыгоден обоим участникам симбиоза. Складывается впечатление, что такой союз был бы невыносим для обеих сторон. Неужели так и начиналась история тесного симбиоза эукариотической клетки с митохондриями? Это не просто маловероятно, это дико. К счастью, есть еще один вариант. До недавнего времени он казался надуманным, но теперь выглядит гораздо убедительнее.
Водородная гипотеза
Поиски предка эукариотической клетки зашли в тупик. Предположение о том, что таким предком могла быть примитивная переходная форма с ядром, но без митохондрий, не было опровергнуто, но становилось все более и более шатким. Все кандидаты на эту роль при ближайшем рассмотрении оказались вовсе не связующим звеном, а вторично упрощенными организмами. Их предки имели митохондрии, а затем митохондрии были утрачены в процессе приспособления к новым экологическим нишам, например к паразитическому образу жизни. Быть эукариотом без митохондрий можно — среди простейших есть тысячи таких примеров, — но быть эукариотом, в родословной которого нет ни одного предка с митохондриями, нельзя. А если так, то нельзя ли предположить, что эукариотическая клетка изначально возникла в результате симбиоза между предками митохондрий (бактериями) и клетками-хозяевами?
Если эукариотическая клетка появилась в результате слияния двух разных клеток, сразу встает вопрос: а каких именно? В учебниках пишут, что клетка-хозяин была примитивной эукариотической клеткой без митохондрий, но это не может быть правдой, если, как мы только что видели, примитивных эукариот без митохондрий не существует. На самом деле, о союзе двух разных бактериальных клеток говорилось еще в теории эндосимбиоза Линн Маргулис, и теперь, когда поиски «недостающего звена» зашли в тупик, ее гипотеза имела хорошие шансы снова привлечь к себе внимание. В любом случае и старая теория Маргулис, и новые рассуждения на эту тему базировались на сходных логических предпосылках: наверное, хозяин производил энергию за счет брожения, как это делают сегодня дрожжи, а митохондрии давали возможность иметь дело с кислородом, то есть предлагали более эффективный способ производства энергии. Кто именно был хозяином, теоретически можно было понять, сравнив генетические последовательности современных эукариот и разных групп бактерий и архей. Как раз в то время появились соответствующие методы секвенирования, однако, как мы только что видели, ответ всех потряс. Гены эукариотических клеток, судя по всему, были ближе всего к генам метаногенов, этих странных обитателей болот и кишок.
Метаногены! Ответ обернулся очередной загадкой. В главе 1 мы говорили, что метаногены живут за счет взаимодействия водорода с углекислым газом, а метан является побочным продуктом их жизнедеятельности. Свободный водород есть только там, где нет кислорода, поэтому метаногены приурочены к аноксическим (бескислородным) местообитаниям. Но на самом деле все еще хуже. Метаногены могут выносить присутствие кислорода, так же как мы можем, задержав дыхание, некоторое время находиться под водой. Проблема в том, что в таких условиях метаногены не могут производить энергию — им приходится «задерживать дыхание» до тех пор, пока они снова не окажутся в аноксическом местообитании, так как их процесс производства энергии возможен только при полном отсутствии кислорода. Если клетка-хозяин действительно была метаногеном, какова же была природа симбиоза? Это серьезный вопрос. С какой стати метаноген должен вступать в союз с бактерией, которой нужен кислород? Современные митохондрии зависят от кислорода, и если это всегда было так, то ни один из участников предполагаемого симбиоза не мог жить на территории другого. Это серьезный парадокс, и он казался неразрешимым.
Однако в 1998 г. на сцену вышел Билл Мартин, с которым мы уже встречались в главе 1. Он и его давний соавтор Миклош Мюллер из Рокфеллеровского университета (Нью-Йорк) опубликовали в Nature статью, в которой представили очень оригинальную водородную гипотезу. Как следует из названия, она имеет мало отношения к кислороду, а вот к водороду — самое непосредственное. Мартин и Мюллер видели разгадку в том, что водород производят в качестве побочного продукта жизнедеятельности странные, похожие на митохондрии органеллы — гидрогеносомы. Они встречаются в основном у примитивных одноклеточных эукариот, включая паразитов, таких как Trichomonas vaginalis (один из несостоявшихся архезоев). Подобно митохондриям, гидрогеносомы производят энергию, но делают это весьма эксцентричным образом, испуская в окружающую среду водород.
Долгое время эволюционное происхождение гидрогеносом было окутано тайной, но ряд их структурных признаков навел Мюллера и других ученых (в особенности стоит упомянуть Мартина Эмбли с коллегами из Музея естественной истории в Лондоне) на мысль о том, что гидрогеносомы родственны митохондриям, вернее, что у них был общий предок. Доказать это было трудно, так как большинство гидрогеносом полностью утратило геном, но тем не менее это удалось сделать[25]. Получается, среди потомков бактерии, вступившей в симбиоз с первой эукариотической клеткой (какой бы она ни была), числятся и митохондрии, и гидрогеносомы. Может быть, говорил Мартин, бактериальный предок митохондрий и гидрогеносом мог выполнять метаболические функции как первых, так и вторых. (В этом вся соль водородной гипотезы.) Таким образом, это была «разносторонняя» бактерия, способная и к кислородному дыханию, и к производству водорода. Мы скоро вернемся к этому вопросу, а пока давайте просто возьмем на заметку вот что: «водородная гипотеза» Мартина и Мюллера утверждает, что именно водородный, а не кислородный метаболизм этого общего предка дал первому эукариотическому организму эволюционное преимущество.
Мартин и Мюллер обратили внимание на один крайне любопытный факт: в клетках эукариот с гидрогеносомами иногда встречаются мелкие метаногены; они когда-то попали внутрь и «загнездились» там. Метаногены тесно прилегают к гидрогеносомам; можно подумать, они с их помощью питаются (рис. 3).
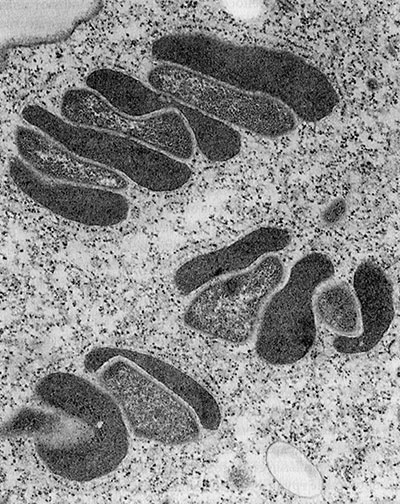
Рис. 3. На этой фотографии видны метаногены (светло-серые) и гидрогеносомы (темно-серые).
Они находятся в цитоплазме значительно более крупной эукариотической клетки — морской инфузории Plagiopyla frontata. Согласно водородной гипотезе, эта тесная метаболическая связь между метаногенами (которым для жизни нужен водород) и производящими водород бактериями (предками митохондрий и гидрогеносом) могла дать начало самой эукариотической клетке: метаногены увеличились в размерах и поглотили производящую водород бактерию
Мартин и Мюллер поняли, что именно так и обстоит дело — эти два организма живут, так сказать, в метаболическом браке. Метаногены уникальны тем, что для производства всех необходимых им органических соединений, а также энергии им нужны углекислый газ и водород, и ничего более. Они присоединяют атомы водорода (H) к молекулам углекислого газа (CO2), и в результате получаются базовые строительные блоки, необходимые для производства углеводов, таких как глюкоза (C6H12O6). Из них они могут строить весь ассортимент нуклеиновых кислот, белков и липидов. Они используют водород и углекислый газ также и для производства энергии, выделяя метан в качестве побочного продукта.
Итак, метаногены отличаются метаболическим хитроумием, и тем не менее они постоянно сталкиваются с очень серьезной проблемой, о которой мы уже говорили в главе 1. Беда в том, что, хотя углекислого газа всегда предостаточно, в любом местообитании, где есть кислород, водорода мало, так как водород и кислород взаимодействуют с образованием воды. Это означает, что с точки зрения метаногена все, что производит хоть немного водорода, — это просто подарок судьбы. А гидрогеносомы — подарок вдвойне, потому что в процессе производства собственной энергии они испускают и водород, и углекислый газ — как раз то, что нужно метаногенам. Что еще важнее, гидрогеносомам не нужен кислород — наоборот, они предпочитают его избегать, и они могут функционировать в условиях дефицита кислорода. Неудивительно, что метаногены присасываются к гидрогеносомам, как поросята к свиноматке! Заслуга Мартина и Мюллера в том, что они поняли, что этот тесный метаболический союз мог быть основой симбиотического происхождения эукариотической клетки.
Билл Мартин утверждает, что гидрогеносомы и митохондрии находятся на разных концах малоизученного спектра разнообразия. К вящему удивлению тех, кто знаком с митохондриями только по учебникам, у многих простых одноклеточных эукариот митохондрии работают в отсутствие кислорода. Вместо него такие «анаэробные» митохондрии используют для «сжигания» пищи другие простые соединения, например нитраты или нитриты. В остальном они очень похожи на «обычные» митохондрии и, несомненно, находятся с ними в родстве. Таким образом, спектр простирается от «аэробных» митохондрий вроде наших, которым нужен кислород, До «анаэробных» митохондрий, предпочитающих другие молекулы, скажем, нитраты, и далее до гидрогеносом, которые функционируют иначе, чем обычные митохондрии, но тем не менее родственны им. Существование такого спектра заставляет задуматься о его происхождении. «Как же выглядел общий предок митохондрий и гидрогеносом?» — спрашивает Мартин.
Этот вопрос очень важен для происхождения эукариот, а значит, всех сложных форм жизни как здесь на Земле, так и во всей Вселенной. Поиск общего предка сводится к выбору одного из двух возможных вариантов. Первая возможность: он был сложной бактерией, гораздой на метаболические ухищрения; некоторые из них ее потомки сохранили, а некоторые утратили в процессе адаптации к новым условиям жизни. Если так, то дальнейшая эволюция этих потомков была не прогрессивной, а регрессивной — они упрощались, переходя к узкой специализации. Вторая возможность: общий предок был простой, дышащей кислородом бактерией, чем-то вроде свободноживущего предка Rickettsia. Ее потомки становились все более разнообразны — они не регрессировали, а прогрессировали. Каждый из этих вариантов позволяет сделать конкретные предсказания. В первом случае, если предковая бактерия была метаболически сложной, она могла передавать специализированные гены, например гены, отвечающие за производство водорода, непосредственно своим потомкам. Любые эукариоты, адаптировавшиеся к производству водорода, могли унаследовать гены от этого общего предка, независимо от того, насколько разнообразными они стали впоследствии. Гидрогеносомы встречаются у разных групп эукариот. Если они получили гены, отвечающие за производство водорода, от общего предка, то эти гены должны быть близко родственны, независимо от того, насколько разнообразными стали содержащие их клетки. С другой стороны, если разнообразные группы и унаследовали изначально простые, использующие кислород митохондрии, то им пришлось придумывать разнообразные формы анаэробного метаболизма независимо каждый раз, когда они оказывались в местообитаниях с низким содержанием кислорода. В случае гидрогеносом, гены, отвечающие за производство водорода, должны были независимо возникать в каждом случае (или перемещаться случайно за счет горизонтального переноса генов), а значит, их эволюционная история выглядит столь же разнообразной, как история клеток, в которых они жили.
Осталось сравнить эти два предположения с действительностью и сделать выбор. Если предок обладал сложным метаболизмом, то все гены, отвечающие за производство водорода, должны быть родственны или, по крайней мере, могут быть родственны. Если же предок обладал простым метаболизмом, то все эти гены должны быть неродственны. Так как же обстоит дело? Окончательного ответа пока нет, но большинство данных, за редкими исключениями, по-видимому, свидетельствуют в пользу первой гипотезы. Несколько исследований, опубликованных в начале нынешнего тысячелетия, говорят о едином происхождении по крайней мере некоторых генов анаэробных митохондрий и гидрогеносом, как и предсказывает водородная гипотеза. Например, фермент, при помощи которого гидрогеносомы образуют водород (пируват: ферредоксин оксидоредуктазы, или ПФОР), был почти наверняка унаследован от общего предка. По-видимому, общее происхождение имеют и мембранный насос, транспортирующий АТФ из митохондрий и гидрогеносом, а также фермент, необходимый для синтеза респираторного железо-серного белка. Эти исследования наводят на мысль, что общий предок действительно был метаболически «разносторонним», то есть мог, в зависимости от обстоятельств, дышать с использованием кислорода или других молекул или производить водород. Такая разносторонность может показаться надуманной, но она действительно существует сегодня у некоторых групп α-протеобактерий, таких как Rhodobacter, которые, соответственно, больше похожи на предковые митохондрии, чем Rickettsia.
Но почему тогда геном Rickettsia так похож на геном современных митохондрий? Мартин и Мюллер предположили, что возникновение параллелей между Rickettsia и митохондриями связано с двумя факторами. Во-первых, Rickettsia являются α-протеобактериями, поэтому их гены, отвечающие за аэробное дыхание, действительно должны быть родственны генам аэробных митохондрий, а также генам других свободноживущих аэробных α-протеобактерий. Другими словами, митохондриальные гены похожи на гены Rickettsia не потому, что они обязательно унаследованы от Rickettsia, а потому, что и Rickettsia и митохондрии унаследовали гены аэробного дыхания от общего предка, который мог быть совсем не похож на Rickettsia. В таком случае встает резонный вопрос: почему, если предок был не похож ни на рикеттсий, ни на митохондрии, их гены в результате стали такими похожими? Этот вопрос подводит нас ко второй гипотезе, выдвинутой Мартином и Мюллером, — гены уподобились друг другу в результате конвергентной эволюции, о которой шла речь в начале первой части книги. Рикеттсии и митохондрии ведут сходный образ жизни в сходных местообитаниях: они производят энергию путем аэробного дыхания внутри других клеток. Их гены испытывают сходное давление естественного отбора, что часто приводит к конвергенции как в плане того, какие именно гены сохраняются, так и в плане последовательности ДНК. Если конвергенция отвечает за сходство, то гены Rickettsia должны быть похожи только на аэробные митохондрии, а не на анаэробные митохондрии, про которые только что шла речь. Если же общий предок очень отличался от Rickettsia (если это была метаболически «разносторонняя» бактерия, такая как Rhodobacter), то параллелей между Rickettsia и этими анаэробными митохондриями быть не должно. И действительно, по большей части их и нет.
|
|
|
