 |


Скелетные мышцы, механизм сокращения мышечного волокна
|
|
|
|
Скелетные мышцы называются также произвольными, поскольку их сокращением и расслаблением можно сознательно управлять, хотя частичное сокращение, обеспечивающее мышечный тонус, регулируется без участия сознания. При исследовании этих мышц на продольных гистологических срезах можно видеть регулярную поперечную исчерченность, отчего мышцы получили название поперечнополосатых. Управление этих мышц осуществляют мотонейроны, входящие в соматическую нервную систему.
Общие принципы сокращения описываются теорией скользящих нитей (рис. 3.1).
Морфологическим субстратом сократимости являются сократительные белки — актин и миозин — это нитевидные белки, расположенные в мышечном волокне параллельно друг другу. При сокращении эти белки скользят друг относительно друга, но сами при этом не укорачиваются.
Взаимное скольжение нитей актина и миозина обусловлено следующим механизмом:
— на нити миозина имеются поперечные мостики, состоящие из шейки и головки;
— на нити актина имеются активные центры, к которым может присоединяться головка миозинового поперечного мостика;
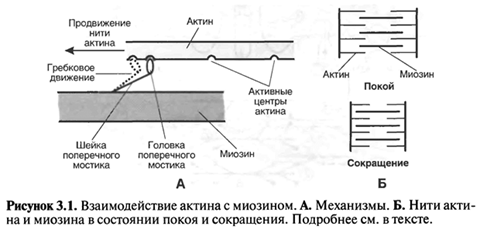 — после присоединения головки поперечного мостика к активному центру актина этот мостик делает вращательное («гребковое») движение, при котором нить актина продвигается вдоль нити миозина.
— после присоединения головки поперечного мостика к активному центру актина этот мостик делает вращательное («гребковое») движение, при котором нить актина продвигается вдоль нити миозина.
В покое актин с миозином не взаимодействуют, и сокращение не происходит. Реакция актина с миозином запускается Са2+. Этот ион действует не непосредственно на актин и миозин, а на те или иные регуляторные белки, разные для поперечнополосатой и гладкой мышц. Взаимодействие актина с миозином требует энергии АТФ.
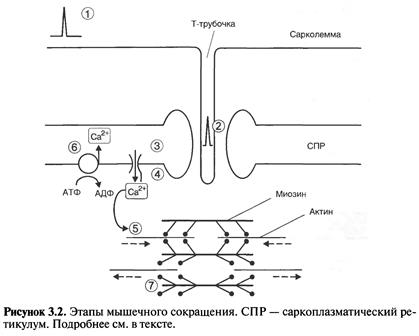 На рис. 3.2 схематично изображено поперечнополосатое мышечное волокно с основными структурами, обеспечивающими его возбуждение и сокращение. К этим структурам относятся:
На рис. 3.2 схематично изображено поперечнополосатое мышечное волокно с основными структурами, обеспечивающими его возбуждение и сокращение. К этим структурам относятся:
|
|
|
- поверхностная мембрана (сарколемма), образующая продольные углубления – Т-трубочки;
- саркоплазматический ретикулум, служащий депо Са2+;
- миофибриллы – пучки параллельных нитей актина и миозина.
Этапы сокращения волокна скелетной мышцы следующие (рис. 3.2)
1. На сарколемме возникает ПД, по своим параметрам и механизмам в основном сходный с ПД нервных клеток.
2. ПД проводится по сарколемме, что приводит к деполяризации Т-трубочек.
3. Деполяризация Т-трубочек приводит к открыванию кальциевых каналов саркоплазматического ретикулума.
4. Из саркоплазматического ретикулума выходит Са2+.
5. Са2+ запускает взаимодействие актина с миозином, происходит их скольжение друг относительно друга и мышца сокращается.
6. Са2+ закачивается обратно в саркоплазматический ретикулум с помощью Са2+-АТФазы (кальциевого насоса).
7. Взаимодействие актина с миозином прекращается – мышца расслабляется.
Са2+действует на сократительные белки (актин и миозин) не непосредственно, а через регуляторные белки. В поперечнополосатой мышце этими белками являются тропонин и тропомиозин. Регуляторные белки, сократительные белки и Са2+ взаимодействуют следующим образом:
— в отсутствие Са2+ активные центры нитей актина прикрыты нитями тропомиозина. С нитями тропомиозина связан тропонин;
— при поступлении к миофибриллам Са2+ этот ион связывается с тропонином;
— взаимодействие Са2+ с тропонином приводит к смещению нитей тропомиозина; активные центры открываются и становятся доступными для присоединения миозиновых поперечных мостиков.
Из этого механизма следуют две важные особенности сокращения одиночного волокна скелетной мышцы:
- сила сокращения пропорциональна концентрации Са2+ в цитоплазме (саркоплазме):чем больше эта концентрация, тем больше открывается активных центров на актине и больше к ним присоединяется миозиновых мостиков.
|
|
|
- сила сокращения одиночного мышечного волокна не зависит от силы раздражителя, то есть подчиняется закону «все или ничего». Это связано с тем, что одиночныйраздражитель независимо от его силы вызывает один ПД, а количество Са2+, выбрасываемое из саркоплазматического ретикулума в ответ на один ПД, всегда одинаково.
|
|
|
