 |


Фосфолипиды: структура, метаболизм и функция
|
|
|
|

Рис. 4—20. Общая структура фосфолипидов.
Природные фосфолипиды являются производными глицерофосфата или сфингозинфосфата, содержащими две жирные кислоты и гидрофильный заместитель, такой, как холин, этаноламин, инозитол или серин. Они служат главными составными частями липопротеинов и клеточных мембран и в большом количестве присутствуют в периферической нервной ткани и головном мозге. Фосфоглицериды обладают общей структурой (рис. 4—20), образующейся путем эстерификации глицерофосфата с двумя длинноцепочечными жирными кислотами и гидрофильным компонентом. Важной особенностью фосфолипидов является их способность взаимодействовать как с гидрофобными, так и с гидрофильными доменами и занимать поверхность раздела между органическими и водными средами. Так, в липопротеинах они выступают в роли связующего звена между белковой частью и транспортируемым нейтральным липидом, а в клеточной мембране они образуют характерный липидный бислой, который ограничивает внутриклеточное пространство.
В плазматической мембране около 60% фосфолипидов приходится на долю фосфатидилхолина, который частично синтезируется в самой мембране путем метилирования фосфатидилинозитола. Фосдатидилинозитол и фосфатидилсерин присутствуют в меньших количествах, причем жирокислотные заместители в разных тканях варьируют по длине и степени насыщенности. Фосдатидилхолины, или лецитины, синтезируются в животных тканях тремя путями. В ходе основного пути, аналогичного таковому для синтеза фосфатидилинозитола, холин фосфорилируется АТФ, образуя фосфохолин. Последний соединяется с ЦТФ, превращаясь в ЦДФ-холин, который реагирует с диацилглицерином в присутствии фосфохолинтрансферазы, образуя фосфатидилхолин. Второй путь синтеза фосфатидилхолина заключается в ацилировании лизофосфатидилхолина (образующегося при гидролизе лецитина фосфолипазой А). Наконец, 3-й путь, имеющий особенное отношение к современным исследованиям гормональных влияний на фосфолипиды мембраны, сводится к последовательному метилированию фосфатидилэтаноламина в клеточной мембране. Эти реакции в схематическом виде представлены на рис. 4—21.
|
|
|
Гормональные влияния на метаболизм фосфолипидов. Стимуляция образования арахидоновой кислоты и синтеза простагландинов.
Многие пептидные гормоны повышают продукцию простагландинов тканью-мишенью, что иногда сопровождается вторичными реакциями сосудов и клеток. Хотя простагландины, по-видимому, не играют роли медиаторов действия пептидных гормонов, но стимуляция этих местно образующихся тканевых гормонов при активации клеток-мишеней составляет важный компонент клеточной реакции на взаимодействие пептидных гормонов с их рецепторами [57]. Простагландины и родственные им простациклины и тромбоксаны быстро синтезируются из полиненасыщенных жирных кислот—предшественников, особенно из арахидоновой кислоты, при стимуляции клеток-мишеней. Ненасыщенные жирные кислоты присутствуют в клетке в виде фосфоглицеридов, которые для того, чтобы превратиться в субстрат для метаболизма в простагландины и другие активные интермедиаты, должны деацилироваться под действием фосфолипаз. На долю арахидоновой кислоты приходится наибольшее количество ненасыщенных жирных кислот в тканевых фосфолипидах; эта кислота подвергается метаболизму в двух основных направлениях, называемых циклооксигеназным и липооксигеназным. Ближайшими продуктами циклооксигеназного пути являются эндоперекиси (ПГG2 и ПГН2), которые превращаются в простагландины (ПГЕ2, ПГF2a и ПГD2) под действием ферментов, носящих общее название простагландинсинтетазы, а также в тромбоксаны (ТКА2 и ТКВ2) и простациклины (ПГI2) под действием соответствующих синтетаз (рис. 4—22). Классификация этих метаболитов эндоперекисей связана со степенью ненасыщенности их жирокислотных предшественников; эйкозатетраеновая кислота превращается в продукты 1-го класса (ПГЕ1, ПГF1a, TKA1 и др.), а арахидоновая кислота—в продукты 2-го класса (ПГЕ2, ПГF2a, ТКА2, ТКВ2 и ПГI2) [58].
|
|
|
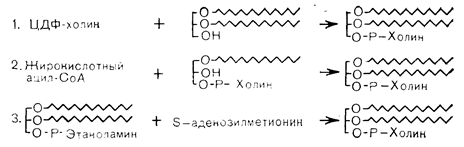
Рис. 4— 21. Пути биосинтеза фосфатидилхолина.
Тромбоксаны и простациклины также образуются при дальнейшем метаболизме эндоперекисей, появляющихся в ходе циклооксигеназного пути; они представляют собой важные регуляторы взаимодействий тромбоцитов с сосудистой стенкой. Простациклин (ПГI2) в больших количествах продуцируется эндотелиальными и гладкомышечными клетками сосудов, где его образование модулируется гидроперекисными метаболитами липооксигеназного пути [59]. ПГI2 является высокоактивным ингибитором агрегации тромбоцитов, а его продукция в эндотелиальных клетках стимулируется тромбином и тормозится липопротеинами низкой плотности. Имеются также данные, свидетельствующие о возможной роли снижения способности гладкомышечных клеток сосудов продуцировать ПГI2 в развитии атеросклероза.
Позднее обнаруженный липооксигеназный путь обусловливает превращение арахидоновой кислоты в оксижирные кислоты, в том числе в гидропероксиэйкозатетраеновую кислоту (ГПЭТЕ), которая затем разрушаясь превращается в оксиэйкозатетраеновую кислоту (ОЭТЕ). Эти метаболиты липооксигеназного пути играют существенную роль в воспалительных реакциях, в том числе хемотаксисе нейтрофилов и других следствиях активации циклооксигеназы в тромбоцитах и продукции ПГI2 в ткани сосудов. Недавно полученные данные свидетельствуют также о том, что липооксигенированные метаболиты арахидоновой кислоты могут опосредовать действие ГнРГ на высвобождение гормонов гипофизарными гонадотрофами [60].

Рис. 4— 22. Образование и метаболизм арахидоновой кислоты. Ненасыщенная жирная кислота, высвобождаемая фосфолипазой А2 из мембранных фосфолипидов, превращается в активные производные в ходе липоксигеназного, циклоксигеназного и простагландинсинтетазного ферментативных процессов.
|
|
|
Любой из перечисленных путей продукции активных метаболитов арахидоновой кислоты зависит от адекватного поступления ненасыщенного жирокислотного предшественника из мембранных фосфолипидов. В настоящее время известно, что многие формы опосредованной рецепторами активации клеток сопровождаются повышением активности связанных с мембранами фосфолипаз, которые катализируют гидролиз эфирных связей в глицерофосфолипидах. Наиболее важна в этом отношении фосфолипаза А2, отщепляющая жирные кислоты во 2-м положении диацилглицерофосфолипидов, которая образует лизофосфолипид и ненасыщенную жирную кислоту, обычно арахидонат. Деацилированный фосфолипид быстро реацилируется за счет переноса активированной СоА жирной кислоты, что легко можно измерить по включению меченой арахидоновой кислоты в фосфолипиды клеток. Этот кругооборот глицерофосфолипида служит источником арахидоновой кислоты для метаболизма по цикло- и липооксигеназному путям (рис. 4—23) и может влиять на проницаемость мембраны и активность других связанных с мембраной ферментов [61].
Активация фосфолипазы А2 зависит от кальция; она происходит при стимуляции клеток надпочечников АКТГ, что приводит к ускорению кругооборота арахидонилфосфатидилинозитола [62]. Этот эффект вызывается также кальциевым ионофором А23187 и может отражать повышение внутриклеточного уровня кальция при действии АКТГ и вторичной стимуляцией фосфолипазы А2 в качестве ранней реакции, сопутствующей АКТГ-рецепторному взаимодействию. Известно, что действие АКТГ на стероидогенез в надпочечниках зависит от кальция, а не только от образования цАМФ. По крайней мере, часть потребностей в кальции для действия АКТГ может быть связана с опосредуемым фосфолипазой A2 кругооборотом мембранных фосфолипидов при активации коры надпочечников.
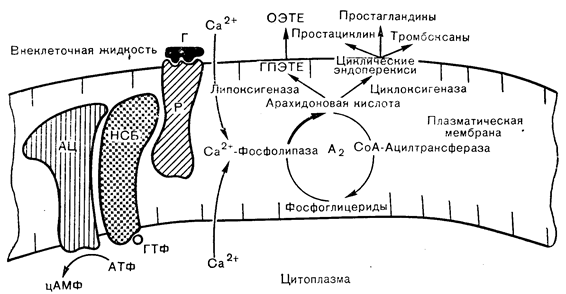
Рис. 4—23. Кругооборот фосфоглицеридов в плазматической мембране с эффектами опосредованного рецепторами (Р) потока кальция на фосфолипазу А2 и продукцию арахидоновой кислоты.
|
|
|
ОЭТЕ — оксиэйкозатетраеновая кислота: Г — гормон; АЦ — аденилатциклаза НСБ — нуклеотид связывающий белок; ГПЭТЕ — гидропероксиэйкозатетраеновая кислота.
Хотя механизм, включающий активацию фосфолипазы, может отражать общее свойство гормонрегулируемых секреторных клеток, при гормональной стимуляции специфических клеток-мишеней меняются и другие этапы метаболизма фосфолипидов. Так, в клетках гранулемы яичника, где ЛГ увеличивает продукцию простагландинов, гормон не повышает образование арахидоновой кислоты, а действует на более поздних этапах, увеличивая активность простагландинсинтетазы [63]. Этот эффект Л Г на синтез простагландинов в граафовом фолликуле (пузырчатый яичниковый фолликул), по-видимому, не опосредует стероидогенного действия гонадотропина, но играет важную роль в развитии овуляции [64].
|
|
|
