 |


Биогеография океанов и морей
|
|
|
|
Экологические области и сообщества организмов океана. Хотя океан представляет собой непрерывную водную оболочку Земли, окружающую материки и острова и обладающую общностью солевого состава, он подразделяется на несколько экологических зон, или областей, с различным набором условий существования организмов.
Прежде всего, различают толщу воды — пелагиаль (от греч. pelagos — море) — область распространения организмов в толще воды или на ее поверхности, и дно океана — бенталь (от греч. benthos — глубина), которая заселена организмами, обитающими на грунте или в грунте (рис.6.1).
Бенталь подразделяется на супралитораль, литораль, сублитораль, батиаль, абиссаль и ультраабиссаль.
Супралитораль — это побережье океана, расположенное выше уровня воды в самый высокий прилив и подверженное только эпизодическому воздействию океанических вод при нагонах и штормах. Здесь обитают как наземные, так и морские организмы.
Литораль (от лат. litoralis — береговой, прибрежный) представляет собой прибрежную часть морского дна, затапливаемую во время приливов и осушаемую при отливах. Она покрывается водой и освобождается от неё дважды в сутки. В зарубежной океанографической литературе нижняя граница литорали иногда определяется глубиной действия волн (40—50 м) или распространения растений (200 м).
Растительный мир литорали состоит в основном из синезеленых, зеленых, бурых и красных водорослей, прикрепленных ко дну. На каменистых грунтах он богаче, чем на илистых. Максимального развития флора литорали достигает в умеренном поясе.
Фауна наиболее разнообразна в тропиках, по направлению к полюсам она становится все беднее. Для животных литораль служит зоной, промежуточной между сушей и морем, и определяет переход от наземного к морскому образу жизни. Многие виды могут в течение того или иного периода оставаться на воздухе, поскольку имеют различные приспособления, защищающие от высыхания во время отлива. Так, морские желуди плотно закрывают раковину, хитоны и морские блюдечки присасываются к камням, актинии и голожаберные моллюски сильно сжимаются и покрываются слизью.
|
|
|
Кроме названных животных на скалах обитают устрицы, мидии, литорины на водорослях и среди них — гидроиды, мшанки, асцидии, в мягких грунтах - многие черви и роющие двустворчатые моллюски. Во время прилива появляются рыбы охотящиеся за обильной пищей.
Сиблитораль является зоной развития жизни на дне океана от уровня воды при самом низком отливе до нижней границы распространения донной растительности. Ей свойственна четкая вертикальная дифференциация водорослей: верхние горизонты занимают зеленые, средние - бурые и самые нижние — красные водоросли. Донная флора (фитобентос) включает несколько видов травянистых цветковых растений: зостеру, посеидонию, талассию и др. Донная фауна (зообентос) сублиторали чрезвычайно обильна и разнообразна. Особенно многочисленны иглокожие, моллюски, черви и ракообразные, а в тропических широтах - кораллы. В сублиторали сосредоточены основные промысловые запасы морских придонных рыб (палтус, камбала и др.).
Батиаль (от греч. bathys — глубокий) занимает материковый склон на глубинах от 200 до 2500-3000 м. Для нее характерно почти полное отсутствие света, значительное давление (до 24,5 МПа), небольшие сезонные колебания температуры и плотности вод, несплошной покров осадков. Растительный мир чрезвычайно беден. Преобладают разнообразные представители зообентоса; много рыб, переходных к абиссальным формам.
Абиссаль (от греч. abyssos — бездонный) — это зона наибольших морских глубин. Области дна океана с глубинами свыше 6000 м выделяются как ультраабиссаль. В воде полностью отсутствует солнечный свет много углекислого газа, температура постоянно низкая (1—2С в полярных областях ниже 0 °С), огромное давление (до 107,8 МПа) и слабая подвижность воды. Грунты полужидкие органогенного или минерального происхождения. Растительный мир представлен только некоторыми бактериальными формами. Ультраабиссальные животные отличаются резко выраженным эндемизмом и древностью. Пищей им служат бактерии, а также «дождь трупов» и оседающий органический детрит. По этой причине все глубоководные животные — детритоеды и хищники. Они слепы или имеют очень развитые глаза, часто телескопические. У многих рыб и головоногих моллюсков есть органы свечения — фотофоры. У некоторых видов светится или вся поверхность тела или ее участки. Окраска животных темная, при отсутствии пигментации тело белесоватое. Скудные запасы пищи — причина малых размеров животных, а низкая температура и обилие углекислого газа — обызвествления скелетов и желеобразности тканей. Уплощенное тело не позволяет животному погружаться в ил, а длинные конечности — ходули, иглы и стебли удерживают его над дном.
|
|
|
Среди глубоководных рыб имеются виды со специальными приспособлениями для ловли добычи (например, рыбы-удильщики с отростками-приманками, снабженными фотофорами). У некоторых рыб (мешкороты и большероты) огромные пасти, часто с острыми, загнутыми вовнутрь длинными зубами (хаумюд и тактостом). Живоглоты, у которых стенки тела и желудок сильно растягиваются, способны заглатывать добычу, по размерам в 2—3 раза большую самих хищников. Интересно, что глубоководные удильщики, живущие на глубинах, где отсутствуют свет и какие-либо сезонные изменения, размножаются весной и летом.
Абиссальные глубины еще мало изучены. Постоянство глубоководной океанической среды на протяжении длительного геологического времени позволило выжить довольно большому количеству древнейших организмов, среди которых рыба латимерия, десятиногие раки, стебельчатые морские лилии и др.
В зависимости от степени освещенности пелагиаль по вертикали делится на три области: эвфотическая (хорошо освещенная), дисфотическая (сумеречная) и а фотическая (света недостаточно для фотосинтеза растений или он полностью отсутствует).
|
|
|
В пелагиали выделяются неритическая и океаническая зоны. Неритическая располагается в пределах шельфа и ограничена глубинами 200 м. Она прерывается пространствами с сильно опресненной и даже пресной водой у устьев больших рек. Многие морские обитатели (ластоногие, пингвины, прибрежные виды китообразных, некоторые виды крабов и раков-отшельников) связаны с берегами. Неритическую зону населяют также животные, личинки которых ведут бентосный образ жизни, а взрослые особи — пелагический, и наоборот. В ней сосредоточены основные районы рыболовства. Океаническая зона включает всю толщу воды, кроме неритической.
В целом в толще воды прослеживаются следующие зоны: п оверхностная (эпипелагиаль), переходная (мезо-пелагиаль) и глубоководная, в которую входят батипелагиаль, абиссопелагиаль и ультраабиссаль.
Толща воды населена представителями планктона (переносимые морскими течениями и волнами), нектона (активно плавающие), плейстона (обитающие на поверхности воды), нейстона (прикрепленные к поверхностной пленке воды снизу или сверху) и гипонейстона (живущие непосредственно под водной поверхностью).
Ареалы морских животных и растений. Ареалы морских организмов различной систематической принадлежности определяются не только термической однородностью водной среды, но и течениями. Среди них встречаются таксономические категории циркумполярного, биполярного и циркумокеанического распространения. Для многих океанических видов характерно амфибореальное (от греч. amphi — по обе стороны) распространение: амфи-атлантическое — вдоль западного и восточного побережий бореальной части Атлантического океана и амфипацифическое — вдоль западного и восточного побережий Тихого океана. В Арктике виды с такими ареалами отсутствуют.
Ряду представителей животных и растений Мирового океана, которые встречаются в бореальной области Северного полушария и в нотальной (аналог бореальной, от греч. notos — юг) Южного полушария, свойственно биполярное распространение. К ним относятся ламинариевые, фукусы (из бурых водорослей), обыкновенные тюлени, котики, несколько видов китов, некоторые акулы, разнообразные рыбы (кильки, сардины) и многочисленные беспозвоночные.
|
|
|
Для объяснения такой дизъюнкции ареалов были выдвинуты различные гипотезы от древнего космополитического распространения до исходного существования в глубине тропических вод. Наиболее вероятная из них принадлежит Л. С. Бергу.
По Л. С. Бергу (1934), биполярность представителей морской фауны и флоры есть следствие охлаждения вод Мирового океана в четвертичный период. В этот период могла осуществиться миграция перечисленных представителей органического мира из одного полушария в другое через тропические и экваториальные воды, особенно там, где расстояние между холодными течениями полушарий невелико (Перуанское, Калифорнийское у западного побережья Америки, Бенгальское и Канарское у западного побережья Африки). Причем северные виды были более активны и сыграли большее значение в возникновении биполярности. Однако в ледниковый период понижение температуры тропических и экваториальных вод было достаточным, чтобы только некоторые представители морской фауны умеренных широт смогли переселиться из одного полушария в другое. Кратковременный характер незначительного охлаждения океанических вод не позволил большинству арктических и антарктических представителей фауны расширить свои ареалы. Последующее потепление климата и вызвало биполярные разрывы ареалов.
Реликты фауны. Как среда, в которой развивается и распространяется жизнь, Мировой океан резко отличается от суши. Отличия связаны в основном с тем, что это водная среда с относительно постоянным солевым составом, мало изменяющимся в пространстве и во времени.
Это свойство обусловило сохранение в Мировом океане представителей древнейших геологических эпох, особенно на больших глубинах с низкой температурой воды. Таковы, например, морские звезды, ежи и стебельчатые морские лилии, обитавшие еще в палеозое. В 1952 г. датское судно «Галатея» подняло с глубины 3950 м моллюска неопилину (Neopilina chulumnae), очень близкого к кембрийскому роду триблидиум. Научной сенсацией стала первая поимка у берегов Южной Африки в 1938 г. латимерии (Latimeria chulumnae), принадлежащей к древним целикантовым рыбам. Палеонтологические остатки целикантовых, от которых произошли наземные позвоночные, неизвестны в слоях моложе мелового возраста. Целикантовые рыбы живут только у островов Анжуан и Большой Комор, где они изредка попадаются местным рыбакам.
|
|
|
Биологическая структура океана и распространение промысловых зон. Жизнь в океане чрезвычайно богата. Морская флора насчитывает около 10 тыс. видов растений: многообразны водоросли, травы, мангровые деревья и кустарники, бактерии, беднее представлены низшие грибы. Повсеместно распространены только бактерии и некоторые низшие грибы. Они участвуют в круговороте веществ, усваивая содержащиеся в воде и донных осадках продукты жизнедеятельности других организмов и будучи пригодными для использования рядом животных. Остальные растительные организмы населяют лишь эвфотическую зону. Водоросли являются основными продуцентами в Мировом океане, производя ежегодно около 100 млрд т органического вещества из общей продукции морской растительности, составляющей 550 млрд т. Среди высших растений обильны травы (около 50 видов) из семейства рдестовых и водокрасовых, образующие настоящие подводные луга на глубинах до. 100 м. Как отмечалось, особенно распространены зостера, посейдония и талассия.
Морская фауна включает около 160 тыс. видов: простейшие (фораминиферы, радиолярии, жгутиковые и инфузории) (10 тыс. видов), губки (5 тыс.), кишечнополостные (9 тыс.), многощетинковые и другие черви (более 7 тыс.), плеченогие и мшанки (свыше 4 тыс.), моллюски (примерно 80 тыс.), ракообразные (более 20 тыс.), иглокожие (6 тыс.), оболочники (1 тыс.), рыбы (16тыс.), черепахи и змеи (50 видов). С океаном и морем связана жизнь более 100 видов млекопитающих, главным образом китообразных и ластоногих, и 240 видов птиц (пингвинов, альбатросов, чаек и др.).
Жизнь в океане распространена довольно неравномерно. Впервые концепция биологической структуры океана была сформулирована В. И. Вернадским в 1926 г. Согласно этой концепции, жизнь в океане сконцентрирована в «пленках» — географических пограничных слоях различного масштаба. Причем ее максимальная концентрация («сгущение») приходится на районы сближения многих пограничных слоев. По мнению ученого, лишь около 2% общей массы океана занято «сгущениями» жизни, а вся остальная содержит рассеянную жизнь. Он выделил четыре постоянных «скопления» жизни: две «пленки» — планктона и донную и два «сгущения» — прибрежное и саргассовое. Прибрежному «сгущению» жизни В. И. Вернадский придавал наибольшее значение, рассматривая его в качестве «области мощной химической активности». Позднее, когда интересы гидробиологии стали фокусироваться на первичной продуктивности и рыбопродуктивности верхнего слоя океана, концепция В. И. Вернадского относительно глобального распределения жизни и ее биохимической активности в океане отошла на второй план.

|
Рис. 2. Наиболее продуктивные зоны прибрежных апвеллингов и системы атмосферного давления на уровне моря (антициклоны) (по Одуму, 1986).
Жирными стрелками показано примерное положение основных течений
В 40—50-е гг. Л. А. Зенкевичем была разработана другая концепция биологической структуры океана. В соответствии с ней все явления в биотической и абиотической средах распространяются в зависимости от положения относительно трех плоскостей симметрии: одной экваториальной и двух меридиональных, проходящих через «середины» океанов и материков. Экваториальная плоскость представляет собой плоскость широтной симметрии, по обе стороны от которой сменяют друг друга четыре зоны специфического температурного режима и связанных с ним биологических особенностей. Экваториальная зона, сравнительно богатая жизнью, сменяется двумя (по одной в северном и южном направлении) субтропическо-тропическими зонами биологического минимума, далее следуют две зоны умеренных широт биологического максимума и две полярные зоны биологического минимума. Меридиональная симметрия выражается в возрастании биомассы планктона (в десятки раз) и бентоса, изменении видового состава организмов и их вертикального распространения от середины океанов к шельфу и неритической зоне. Нарушения меридиональной симметрии (асимметрия) закономерны и связаны с наличием океанических течений и других явлений в водной среде. Концепция Л. А. Зенкевича подтвердила значение береговой границы раздела и прибрежного сгущения жизни.
Концепции биологической структуры океана, предложенные В. И. Вернадским и Л. А. Зенкевичем, не исключают, а дополняют друг друга. Первая из них, подчеркивающая биохимическую активность организмов, оказалась весьма важной в связи с загрязнением водной среды. Вторая больше ориентирована на изучение распределения жизни в океанах, его видового разнообразия и биологической продуктивности.
Жизнь в Мировом океане сконцентрирована около берегов, где наиболее благоприятны условия питания живых организмов. По сравнению с прибрежными водами и лиманами большая часть открытого океана представляет собой «пустыню». Однако и в прибрежных районах жизнь распространена довольно неравномерно. Максимальная ее концентрация приурочена к коралловым рифам и эстуариям.
Большую роль в распределении жизни в океане играет процесс, названный апвеллингом. Он происходит там, где ветры постоянно отгоняют поверхностную воду от крутого берегового склона (рис. 2). В результате на поверхность поднимается холодная глубинная вода, богатая биогенными элементами. Как правило, апвеллинги расположены у западных берегов континентов, которые нередко заняты пустынями. Образование пустынь связано с тем, что ветры, формирующие апвеллинги, дуют большей частью с суши, унося влагу. Во многих случаях только частые морские туманы поддерживают развитие своеобразной растительности (вельвичия удивительная в пустыне Намиб).
В противоположность пустынным берегам зоны апвеллинга характеризуются колоссальными популяциями рыб и птиц и являются наиболее продуктивными океаническими областями. В коротких пищевых цепях, что также свойственно зонам апвеллинга, доминируют диатомовые водоросли и сельдевые рыбы. Большие популяции морских птиц на берегах и островах откладывают бесчисленные тонны гуано, богатого нитратом и фосфатом. Осадки на морском дне также содержат много фосфата и органических веществ. Изменение направления ветра, когда в бескислородной водной среде происходит «цветение» ядовитых водорослей динофлагеллат, вызывает массовую гибель рыбы. Наиболее часто подобные катастрофы происходят в зоне перуанского апвеллинга (Одум, 1986).
Биогеографическое районирование. При биогеографическом районировании Мирового океана, которое носит зональный характер, А. Г. Воронов (1987) использовал схему зоогеографического районирования, предложенную В.Г. Гептнером (1936). В результате были выделены семь биогеографических областей: Арктическая, Бореально- Тихоокеанская, Бореально-Атлантическая, Тропико-Индо-Тихоокеаническая, Тропико-Атлантическая, Нотально-Антарктическая (Субантарктическая) и Антарктическая (рис. 3).
Эти области являются общими для неритической и океанической водной среды, но не учитывают своеобразие жизни в бентали и пелагиали, других экологических зонах Мирового океана. В частности, в зоогеографическом районировании Мирового океана по донной фауне материковая отмель разделена на три царства: южных и умеренных морей Северного полушария, тропическое, холодных и умеренных морей Южного полушария, а абиссаль на три области: Тихоокеанско-Северо-Индийскую, Атлантическую и Антарктическую.
Арктическая область. Ее южная граница между Северной Америкой и Европой проходит от острова Ньюфаундленд в направлении архипелага Шпицберген к северо-западной оконечности Кольского полуострова, между Азией Северной Америкой--по северной части Берингова моря. В области постоянно низкая температура воды (около 0°С), соленость ниже средней океана (результат таяния льда и выноса пресных вод реками). Зимой и летом преобладающая часть акватории покрыта ледовым панцирем, встречаются полыньи и разводья.
Растительность более холодных вод отличается преобладанием бурых водорослей, а теплых -- зеленых. Красные водоросли представлены сравнительно равномерно.
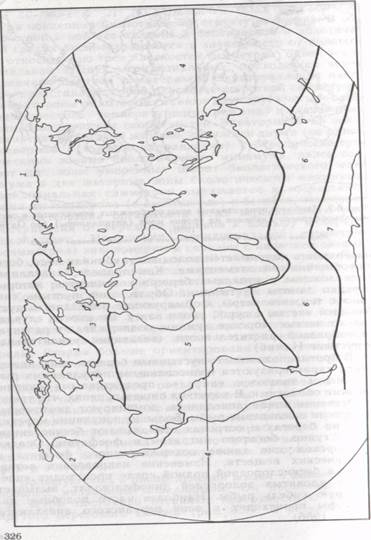
Рис.3.Биогеографическое районирование Мирового океана (по Воронову, 1987): Арктическая; 2 — Нереально-Тихоокеанская; 3 — Бореально-Атлантическая; Тропико-Индо-Тихоокеанская; 5— Тропико-Атлантическая; 6— Субантарктическая; 7 — Антарктическая
Летом у кромки тающих льдов образуется массовое скопление планктона, которое определяет богатое разнообразие водных обитателей от крылоногих моллюсков лиматин до нарвалов и белух (из китообразных), а также чаек и чистиков. Среди чаек эндемична белая, из чистиков характерен люрик. Гренландский кит, обильный раньше, почти полностью истреблен. Из тюленей обычны нерпа, морской заяц, хохлач и морж. Круглый год по льдам бродит белый медведь. Из рыб характерны сайка и навага. Из беспозвоночных много бокоплавов и равноногих, которые достигают здесь наибольшего разнообразия.
Бореально-Тихоокеанская (Бореально-Пацифическая) и Бореально-Атлантическая области. Эти области имеют значительное сходство. Им свойственны резкие сезонные колебания температуры воды — от 3 до 15° и более. Большинство организмов, обитающих здесь, эвритермны. По световому режиму бореальные области океана отличаются от Арктической круглогодичной ежесуточной сменой дня и ночи, что позволяет растительным организмам вегетировать большую часть года.
Границы областей неодинаковы для организмов, обитающих в пелагиали на разных глубинах. Если в районировании учитывать сублиторальную фауну, то южная граница пройдет между 30 и 40° с. ш. и почти совпадет со среднегодовой изотермой 15°С. Система теплых и холодных течений у восточных берегов, как Северной Америки, так и Азии клинообразно суживает акваторию областей.
По видовому разнообразию бореальные флора и фауна значительно превосходят арктическую, но уступают тропической. Наибольшее обилие организмов приурочено к литорали и сублиторали.
Для Бореально-Тихоокеанской области особенно характерны бурые водоросли (макроцистис и нереоцистис) Среди моллюсков обычны устрицы, мидии, морские гребешки, тихоокеанский кальмар и осьминог Дофлейна. Ракообразные представлены вислоногими (каланус тихи океанский) и десятиногими раками (камчатский краб, креветки чилимы). Из иглокожих наиболее распространен трепанг. Рыбы (кета, горбуша, чавыча, нерка, иваси и др.) имеют большое промысловое значение. Из птиц особенно много чистиковых. Разнообразны млекопитающие: морской котик, сивуч, калан, японский кит, серый кит, дельфин, белокрылая морская свинья.
В Бореально-Атлантической области обильны бурые водоросли (ламинария, алярия, фукусы) и красные (анфельция). Ракообразные представлены вислоногими раками, креветками, лангустами, омарами, рыбы -- треской, пикшей, сайдой, кефалью, камбалой и зубаткой. Среди птиц много чаек, чистиков, кайр, гагарок, ластоногих - хохлачей, серых и гренландских тюленей, китообразных -- гринд.
Некоторые представители фауны (серый дельфин и обыкновенная морская свинья, обыкновенный тюлень, чистики, сельдевая акула, морские сельди, треска и др.) имеют амфибореальное распространение у берегов Евразии и Северной Америки. Они отсутствуют в морях, омывающих северные побережья этих континентов. Л.С. Берг объяснил эту разорванность ареалов тем, что температура воды полярных морей в плиоцене была значительно более высокой и многие виды морских животных могли проникнуть из Северной Атлантики через полярные моря, омывающие северное побережье Евразии, в северную часть Тихого океана, и наоборот. Наступившее в антропогене похолодание вызвало резкое понижение температуры в полярных морях, и многие виды вымерли. В более южных широтах понижение температуры воды было не столь значительным, что способствовало сохранению формирующейся амфибореальной фауны.
Тропика-Атлантическая и Тропико-Индо-Тихоокеанская области. Для них также характерны общие черты природных условий и биоты. Прежде всего, постоянно высокая температура поверхностных слоев воды (выше200С) с незначительными годовыми колебаниями (менее 2°), а также контрастные температурные различия между поверхностными и глубинными горизонтами. Северная граница этих областей совпадает с годовой изотермой воды 15 °С, а южная (в Южном полушарии) 170С.
Только в этих областях распространены сообщества мангров и коралловых рифов. Из водорослей следует отметить саргассовые (саргассум и турбинария).
В водах тропических областей обитает подавляющее большинство видов планктонных фораминифер, крылоногих и киленогих моллюсков, сифонофор, кольчатых червей, сальп и аппендикулярий. В тропических областях обычны крупные акулы, летучие рыбы, меч-рыбы, парусники, скаты, морские черепахи, морские змеи, лангусты, жемчужницы. С тропическими морями связаны птицы фаэтоны и фрегаты, большую часть жизни, проводящие в поисках пищи над водной поверхностью. Из млекопитающих обычны белобрюхие тюлени, кашалоты, дюгони и лимантины. Хотя в целом фауна тропических областей океана отличается большим разнообразием, она имеет значительно меньшую, чем в других поясах, численность отдельных видов. Только в районах апвеллингов наблюдаются массовые скопления животных.
Нотально-Антарктическая область. По условиям существования жизни эта область сходна с бореальными. Для нее характерны те же резкие сезонные колебания температуры, подъемы глубинных вод, обогащенных биогенными веществами, и обилие жизни.
Кроме биполярно распространенных представителей флоры и фауны (бурые водоросли, несколько видов китов, котики, обыкновенные тюлени, кильки, сардины и др.) следует отметить: из млекопитающих — гривистого сивуча, южного котика, южного и карликового китов, морского слона и морского леопарда, из рыб представителей семейства нототениевых, а птиц — королевского альбатроса.
Антарктическая область. Как и Арктической, ей свойственны постоянно низкие температуры воды и развитие ледового покрова. Кроме того, условия существования жизни ограничены многочисленными айсбергами, которые как бы перепахивая приливно-отливную полосу, уменьшают обилие жизни в литорали.
Антарктические воды более благоприятны для живых организмов, чем суша, и по этой причине их фауна несравненно разнообразнее. Морские беспозвоночные криль (планктонные ракообразные из семейства Euphausiidae) летом в поверхностных слоях воды образуют огромные скопления, служащие кормом для ряда видов рыб, птиц и млекопитающих. Из рыб распространено семейство белокровных щук. Летом многочисленны буревестники и поморники, нередко встречаются крачки, альбатросы и качурки. Наиболее типичные представители области -- пингвины. У берегов Антарктиды, близ островов и среди дрейфующих льдов обитают настоящие тюлени (Уэдделла, Росса, крабоед, морской леопард морской слон). Довольно многочислен морской котик. Массовое скопление криля привлекает большие стада китов (синего, финвала, горбача, сейвала, полосатика и др.). Встречаются кашалоты, касатки и бутылконосы.
Своеобразна донная фауна Антарктической области. Обильны губки и иглокожие. Масса медуз достигает 156 кг.
К сожалению, флора и фауна Мирового океана изначительной мере испытали на себе разрушительное воздействие антропогенного фактора. Не только уменьшилась численность их представителей, но и полностью уничтожены, отдельные виды, загрязнены нефтью, нефтепродуктами, бытовыми стоками и разнообразными токсичными веществами промышленного происхождения воды.
Биомы островов
Биоты островов морей и океанов чрезвычайно разнообразны. Тесно связанные с условиями образования самих островов, они, однако, не имеют ни одной черты, которая была бы свойственна всем островам без исключения.
Разнообразие биоты острова зависит от его происхождения, возраста, размеров, удаленности от материка и свойственных ему природных условий.
По своему происхождению острова в морях и океанах разделяются на две основные группы – материковые и океанические. Материковые представляют собой отделившуюся в ту или иную геологическую эпоху часть континента. Они выступают над уровнем воды в пределах подводной окраины материков и сложены обычно коренными породами (Великобритания, Гренландия, Мадагаскар, Новая Зеландия, Новая Каледония, Гавайские и др.). Мелкие могут образовываться в результате аккумулятивной деятельности волн и прибоя. Океанические острова распространены в пределах ложа океанов и на срединно-океанических хребтах. К ним относятся коралловые (атоллы и рифы) и вулканические острова (Галапагосские, Кергелен, Маскаренские, Канарские, Пасхи и др.).
Между этими двумя группами островов, материковых и океанических, встречаются промежуточные формы -- геосинклинальные острова, или острова переходной зоны и островных дуг. Они отличаются наибольшим разнообразием природных условий и имеют более сложную структуру, ландшафтов -- от равнинных до горных территорий. Как правило, все геосинклинальные острова формируются в архипелаги или образуют островные дуги (Большие Зондские, Филиппинские, Японские и др.).
По сложности экосистем острова могут быть низкими, включая биогенные (атоллы, рифы и мангровые) и шельфовые, и высокими (вулканические, геосинклинальные материкового склона, или шельфовые). Высокие острова, в свою очередь, могут иметь слабо или хорошо выраженную высотную поясность.
Условия для формирования биоты на материковых и океанических островах совершенно различны. Материковые острова отделились от континентов с тем видовым разнообразием растительного покрова и животного населения, которое было присуще этому участку континента. Океанические острова заселялись только организмами, способными преодолеть большие водные пространства. В первом случае тип формирования биоты может быть определен как реликтовый, во втором--как иммиграционньй.
На островах материкового происхождения биота со временем постепенно обедняется за счет вымирания части форм и даже видов. Вымирание вида можно объяснить его малой численностью в момент отделения острова от материка, не обеспечивающей длительное существование в условиях изоляции. Причем гибель видов, унаследованных островом, не компенсируется появлением иммигрантов, которые попадают на остров, как правило, и ограниченном числе особей и не всегда выживают.
О постепенном вымирании видов на островах можно судить по тому, что небольшие по площади острова материкового происхождения в отличие от крупных имеют почти чисто океаническую фауну. Так, среди островов Пирл на большом острове Рей обитают около 1/3 континентальных видов, а на маленьком острове Кондатора --лишь 1/10 (Воронов, 1987). Если два острова разной величины одновременно отделились от материка, то исходная биота может почти полностью сохраниться на большем из них и полностью или частично исчезнуть на меньшем.
Изоляция и удаленность от материка определяют высокий эндемизм флоры и фауны. Чем древнее остров, тем больше эндемичных видов и форм. Например, видовой эндемизм флоры Новой Зеландии, Новой Каледонии, Гавайских островов достигает 70—80%. При меньшей отдаленности от материковой суши (острова Великобритания, Ирландия, Японские, Шри-Ланка) эндемизм менее выражен.
На островах нередко в облике тех или иных групп животных наблюдаются отклонения. Например, крупные млекопитающие обычно мельче, чем на материке (пони, филиппинский буйвол и др.). У птиц и пресмыкающихся, наоборот, выражен островной гигантизм (вараны на острове Комодо, черепахи на Галапагосских островах). Причина этого явления пока не выяснена.
Нередко для островов характерны нелетающие птицы и насекомые. Происхождение нелетающих птиц связано с отсутствием на островах млекопитающих, которые могли бы их истребить. В отборе нелетающих насекомых важную роль сыграл их снос ветром и ураганами в океан. Для многих видов летающих насекомых устанавливается равновесие между числом особей, унесенных ветром, и числом особей, приносимых на остров, при условии, что остров входит в архипелаг.
В своем распределении биоценозы островов подчиняются тем же зональным закономерностям, что и сообщества континентов. Однако их структура и энергетические связи при меньшем по объему видовом составе проще. Только на больших островах с горными системами (Мадагаскар, Новая Зеландия, Куба, Великобритания и др.) растительный покров и животное население не менее сложны, чем на сопредельных материках. На островах с упрощенной ландшафтной характеристикой сообщества более однотипны, а обеднение видового состава более значительно.
На островах океанического происхождения сообщества возникают на продуктах вулканической деятельности или коралловых известняках. Флора и фауна этих островов полностью иммиграционная, однако, по возрасту, она может оказаться старше самих островов. Проникновение видов с материка на тот или иной остров облегчается по так называемым «мостам суши» и по цепочке островов вулканического или иного происхождения. На некоторых из этих островов вид мог иметь временное пристанище и переселиться на вновь образовавшийся остров в качестве реликта, возникшего в результате вымирания на соседних участках суши. Таким образом, не исключается возможность элемента реликтового происхождения флоры и фауны на островах океанического происхождения.
Перенос организмов через водное пространство по отношению к каждой особи носит случайный характер. При длительном существовании миграционного процесса нот перенос приобретает определенную статистическую вероятность. Так, при пересечении пространства шириной 100 миль выживает лишь одна особь из тысячи, следующих 100 миль — опять одна особь из тысячи и т.д. Шанс достигнуть остров, расположенный в 200 милях от источника миграции, имеет одна особь из миллиона.
Наиболее распространенными способами заселения островов организмами являются гидрохория (морские течения), анемохория (ветры, штормы и ураганы) и зоохория (перенос с помощью птиц). В заселении островов растениями и животными большую роль играет человек (антропохория).
Активно заселять острова могут в основном птицы, однако этот процесс сдерживается «гнездовым консерватизмом» самих птиц. Взрослые растения, прибитые волнами к берегу, как правило, редко приживаются. Выживают эпифиты, находящиеся на стволах. Ветром на большие расстояния переносятся споры и легкие семена, в связи с чем, например, папоротники на островах имеют широкое распространение. Насекомые в целом плохо переносят пребывание в соленой воде и заселяют острова в случае заноса их ветром или птицами. Гибнут в морской воде амфибии, рептилии и пресноводные рыбы. Из рептилий на островах распространены только гекконы и сцинки. В конечном итоге гибель в морской воде приводит к довольно бедному видовому составу этих групп животных. Так, в Южной Америке много земноводных, а на соседних Галапагосах нет ни лягушек, ни саламандр, отсутствуют виды птиц, характерные для материка, а из наземных млекопитающих представлены по одному роду летучих мышей, крыс и грызун, напоминающий хомяка.
Большое значение в заселении острова организмами, переносимыми ветром и в меньшей степени водой, имеет его «ловчий угол»-- расположение острова по отношению к потоку мигрантов. Так, если остров расположен перпендикулярно к потоку мигрантов, то вероятность того, что мигрант попадет на остров, больше.
Флора островов океанического происхождения отличается малочисленностью видового состава. В их фауне отсутствуют млекопитающие, земноводные и змеи.
Процесс видообразования на океанических островах протекает быстрее, чем на материковых и континентах, так как на океанических островах образуется, как правило, малочисленная популяция какого-либо вида с обедненным генофондом. К тому же эта локальная малочисленная популяция оказывается в географической изоляции. В результате на разных островах возникают различия в наборе генов одного и того же вида, приводящие к возникновению на каждом острове внутривидовых форм, а впоследствии и видов. Большое значение при этом имеет и незаполненность экологических ниш.
В качестве примера, достоверно подтверждающего многообразие видообразования на островах океанического происхождения, можно привести вьюрков, обитающих на Галапагосах.
Ч. Дарвин в своем дневнике «Путешествие натуралиста вокруг света на корабле „Бигль"» отмечал, что естественная история этих островов в высшей степени интересна и вполне заслуживает внимания. Большинство обитающих здесь организмов являются аборигенами, в других местах не встречающимися. Этот «спутник Америки», отделенный от материка пространством открытого океана в 500—600 миль, получил с континента нескольких случайных колонистов.
Очевидно, на Галапагосы попал один вид вьюрка, питающийся свойственной ему пищей. Вьюрки, которым не хватало корма, погибали или переходили на другой корм. Так, они научились ловить насекомых, вытаскивать личинки из щелей в стволах деревьев, разгрызать орешки, питаться листьями растений и т. д. В конечном итоге естественный отбор привел к образованию специализированных форм, которые заполнили на острове все экологические ниши, предназначенные для певчих птиц. На Галапагосах существуют 3 рода, 13 видов и 37 островных форм вьюрков: большой, средний и малый земляные, кактусовый, толстоклювый древесный, дятловый, попугайный, кокосовый, мангровый, славковый и др. Интересно, что дя
|
|
|
