 |


Восприятие цвета с позиции векторной модели обработки информации
|
|
|
|
Анализатор цвета включает рецепторный и нейронный уровни сетчатки, ЛКТ таламуса и различные зоны коры. На уровне рецепторов падающие на сетчатку излучения видимого спектра у человека преобразуются в реакции трех типов колбочек, содержащих
пигменты с максимумом поглощения квантов в коротковолновой, средневолновой и длинноволновой частях видимого спектра. Реакция колбочки пропорциональна логарифму интенсивности стимула. В сетчатке и ЛКТ существуют цветооппонентные нейроны, противоположно реагирующие на пары цветовых стимулов (красный-зеленый и желтый—синий). Их часто обозначают первыми буквами от английских слов: +К-С; -К+0; +У-В; -У+В. Различные комбинации возбуждений колбочек вызывают разные реакции оппонен-тных нейронов. Сигналы от них достигают цветочувствительных нейронов коры.
Свойства цветовых нейронов различных зон коры обезьяны подробно изучены английским исследователем С. Зеки. В стриар-ной коре (VI) он нашел два типа цветочувствительных нейронов:
цветооппонентные нейроны и нейроны с избирательной чувствительностью к определенному и узкому диапазону длин волн. Цветооппонентные клетки по своим свойствам совпадают с аналогичными нейронами сетчатки и ЛКТ. Исключение составляет группа нейронов с двойной оппонентностью, у которых и центр и периферия рецептивного поля приобрели способность отвечать по оп-понентному типу. Нейроны в VI, избирательно реагирующие на узкий диапазон спектра, меняют свои реакции на цветовые стимулы с изменением общего освещения. По данным С. Зеки, клетки этих двух типов составляют большинство цветочувствительных нейронов в VI, и их очень мало в экстрастриарной коре (У4), основная часть которой заполнена третьим типом нейронов — цветокодирующими клетками, или детекторами цвета. Они реагируют избирательно на определенный цвет поверхности независимо от спектрального состава освещения. Однако важным условием реакции нейронов-детекторов цвета является наличие освещения, содержащего излучение всех трех областей спектра: коротко-, средне- и длинноволновой. При монохроматическом освещении нейрон-детектор цвета не реагирует на оптимальный для него цвет. Наиболее часто встречаются клетки с чувствительностью к полосе 480 им (к синему), 500 нм (зеленому), 620 нм (красному) и реже к 550—570 нм (желтому). Клетки поля У4 организованы в вертикальные колонки. Колонки, детектирующие разные цвета (красные, синие, зеленые, белые и др.), чередуются. Границы цветовых колонок не совпадают с границами колонок, организованных по другим признакам (ориентации, глазодоминантности). В поле VI цветоселективные нейроны относятся к аконстантным нейронам, их реакция зависит от общего освещения. В поле У4 представлены цветоконстантные нейроны — собственно детекторы цвета, которые выделяют цвет объекта независимо от условий освещения.
|
|
|
Восприятие цвета определяется не только хроматической (цве-точувствительной) системой зрительного анализатора, но и вкладом ахроматической системы. Ахроматические нейроны образуют локальный анализатор, детектирующий интенсивность стимулов. Первые сведения об этой системе можно найти в работах Р. Юнга, показавшего, что яркость и темнота в нервной системе кодируются двумя независимо работающими каналами: нейронами В (Ьп@Ь1пе5з), измеряющими яркость, и нейронами В (оагкпезз), оценивающими темноту. Существование нейронов-детекторов интенсивности света было подтверждено позже, когда в зрительной коре кролика были найдены клетки, селективно реагирующие на очень узкий диапазон интенсивности света.
|
|
|
Как можно представить механизм восприятия цвета? С позиции детекторной теории основу для субъективных различий стимулов следует искать в различии реакций нейронов-детекторов. Отсюда следует, что субъективные различия между цветовыми стимулами пропорциональны расстояниям между представляющими их нейронами-детекторами на нейронных картах признаков, а матрицу субъективных различий, полученных в психофизическом эксперименте, можно интерпретировать как матрицу «расстояний» между детекторами (Соколов Е.Н., 1993). Обработка матрицы субъективных различий, полученной при попарном предъявлении цветовых стимулов, факторным анализом или методом многомерного шкалирования, позволяет построить геометрическую модель субъективных оценок, которая одновременно является моделью нейронального отображения цветовых стимулов на карте детекторов. В такой модели цветовые стимулы представлены точками, а расстояния между ними соответствуют расстояниям между нейронами-детекторами.
Метод построения субъективного пространства позволяет определить его минимальную мерность и выделить основные субсистемы (факторы), на основе которых субъект выносит суждение о различии двух цветовых стимулов. Следующий шаг предполагает интерпретацию выделенных факторов в понятиях нейрональных единиц, образующих нейронные сети детекторов.
Для построения перцептивного пространс^ва цвета у человека на основе оценок субъективных различий были использованы цветовые стимулы разной длины волны и интенсивности (1гтапоу сп.а., 5око1оу Е.М., 1992). Цвета предъявлялись парами с коротким интервалом между стимулами в паре. После предъявления каждой пары испытуемый должен был оценивать различие между ними в баллах от 0 (полностью идентичные) до 9 (максимально различные). Полученные оценки субъективных различий
сводили в матрицу, которую обрабатывали методом многомерного шкалирования (ММШ). Было получено четырехмерное эвклидово пространство — гиперсфера, в которой все множество цветовых стимулов размещалось в ее тонком поверхностном слое (при вариативности ее радиуса не более 10%). Проекция стимулов на плоскость осей Х,Х^ образует круг. Полюса осей Х,Х^ представлены парами стимулов: соответственно красным—зеленым и желтым—синим. Белый цвет (ахроматический) расположен в центре круга. В плоскости Х,Х^ отражен цветовой тон, который для каждого стимула определяется через угол, образованный радиусом к данному стимулу. Проекция точек на плоскость ХдХ^ позволяет интерпретировать ее оси как биполярную яркостную и монополярную темновую составляющие цвета соответственно. Белый цвет расположен вблизи положительного полюса оси Хд, а наиболее темный, например красный, — ближе к отрицательному полюсу оси Хд. Цвета промежуточных значений сдвинуты к положительному полюсу Х„. По координатам цветовых стимулов в плоскости ХдХ^ можно измерить ахроматическую составляющую цветового зрения. Углы на плоскости ХдХ^, образованные радиусами к стимулам, характеризуют их субъективную яркость (светлоту).
|
|
|
Координаты по зелено-красной и желто-синей осям цветового субъективного пространства человека соответствуют реакциям зелено-красных и сине-желтых оппонентных нейронов ЛКТ обезьяны при предъявлении ей тех же цветовых стимулов (с идентичными длинами волн). Координаты двух ахроматических осей цветового перцептивного пространства соответствуют реакциям ахроматических нейронов: яркостных (нейроны В) и темновых (нейроны О). Вычисление обобщенной хроматической оси (для Х^Хд) по формуле (X^+X;)1/2 и обобщенной ахроматической оси для ХдХ^ по формуле (Х^+Х^)172 преобразует четырехмерное пространство в плоскость, в которой цветовые стимулы располагаются на ограниченном участке дуги первого квадранта в зависимости от их насыщенности. Углы в этой плоскости, образованные радиусами, характеризуют стимулы по их насыщенности. С увеличением угла насыщенность стимула уменьшается.
В представленной модели цветового зрения органически соединяются нейронный механизм кодирования цвета с психологическими характеристиками цвета: цветовым тоном, светлотой (субъективной яркостью) и насыщенностью, представленными в угловых характеристиках на плоскостях X^Xу ХдХ^ и в плоскости обобщенных хроматической и ахроматической осей.
|
|
|
Перцептивные цветовые пространства, построенные для животных (трихроматов) — обезьяны (макаки), рыбы (карпа), об-
наруживают их принципиальное сходство с субъективным цветовым пространством человека (Латанов А.В. и др., 1997). В опытах на животных был использован метод измерения вероятности инструментальных двигательных реакций при выработке цветовых диф-ференцировок. После обучения цветовой стимул (как условный сигнал) характеризуется вектором вероятностей реакций, отражающим вероятность появления условной реакции как на условный, так и на дифференцировочные стимулы. Последовательно проводя серии с подкреплением разных цветов, можно для каждого цвета составить матрицу вероятностей условных рефлексов и на ее основе вычислить корреляционную матрицу для последующей обработки факторным анализом. Перцептивные цветовые пространства этих животных, так же как и человека, представляют гиперсферу, оси которой соответствуют четырем каналам кодирования цвета:
красно-зеленому, желто-синему, яркостному и темновому (рис. 10, 11). Три угла цветовой гиперсферы соответствуют трем субъективным характеристикам цветовых стимулов: цветовому тону, светлоте и насыщенности.
Серьезным подтверждением правомерности применения психофизического подхода для раскрытия нейронных механизмов восприятия явились опыты, выполненные на лягушке (трихромате) с регистрацией электроретинограммы (волны «Ь») на смену цветовых стимулов (Зимачев М.М. и др., 1991). Был использован метод замены одного (тестового) цвета на другой (референтный) при меняющейся яркости тестового цвета. Каждому стимулу соответствует вектор, представляющий величины волны «Ь&, полученные при замене данного стимула на каждый другой. Из этих векторов составляли матрицу смешений, которую обрабатывали ММШ. Перцептивное цветовое пространство, полученное по волне «Ь» сетчатки лягушки, обнаружило большое сходство с перцептивным пространством человека, построенным на основе субъективных оценок, перцептивными пространствами обезьяны и карпа при измерении у них вероятностей условных двигательных реакций. Особенностью, отличающей перцептивное цветовое пространство лягушки, была меньшая степень дифференциации цветовых стимулов, что выражалось в их группировке вокруг основных цветов:
красного, зеленого, синего, желтого и белого. Авторы предполагают, что у лягушки отсутствует механизм различения насыщенности цвета, т.е. хроматические и ахроматические системы не образуют нового качества, представленного углом в плоскости обобщенных хроматической и ахроматической осей-
|
|
|
Правомерность четырехмерной сферической модели цветораз-личения подтверждается опытами с предъявлением стимулов, часть

|

|
й.- / „ - Е "5 5 \ а


|

|
параметров которых редуцирована. В психофизических опытах на человеке, а также в поведенческих экспериментах на животных показано, что в тех случаях, когда предъявляемые стимулы различаются только по яркости, их распознавание осуществляется при участии только двух ахроматических каналов. Выявленные в этих опытах перцептивные пространства яркости для человека и животных двумерны. При различении равноярких стимулов разного спектрального состава перцептивное пространство трехмерно. Это было продемонстрировано в психофизических исследованиях на человеке, а также на обезьянах, когда пространство строилось из матрицы смешения, составленной из вероятностей инструментальных ответов обезьяны при выработке дифференцировок по цветовому тону. Третья координата такого пространства, соответствующая светлоте, отражает интегрированный вклад двух ахроматических каналов. Таким образом, трехмерное перцептивное пространство равноярких цветовых стимулов и двумерное пространство яркости представляют собой подпространства общего четырехмерного цветового перцептивного пространства.
Психофизический метод изучения восприятия цвета, включающий построение геометрической модели перцептивного пространства цвета по матрице речевых или поведенческих реакций, дает возможность охарактеризовать цвет в векторных понятиях. Таким образом, субъективное восприятие каждого цвета представлено четырехкомпонентным вектором, определяемым вкладами по четырем осям гиперсферы.
С позиции векторной психофизиологии все детекторы цвета представлены точками на сферической поверхности (карте детекторов) четырехмерного пространства. Сферическая карта цветовых детекторов имеет дискретную структуру. Порог различения цветов определяется расстояниями между соседними детекторами. Величина порога зависит от плотности детекторов на детекторной карте. При одинаковой их плотности порог различения одинаков на разных участках детекторной карты. Изменение цвета обнаруживается при смещении максимума возбуждения с одного детектора на соседний.
Нейрон-детектор представляет вершину иерархически организованной нейронной сети (или ансамбля нейронов). Ансамбль нейронов — это группа нейронов, имеющая общий для них вход и конвергирующая на более высоком уровне на одном или параллельно на группе нейронов.
Другими словами, ансамбль нейронов, конвергирующих на нейроне-детекторе, образует его нейронную сеть. В ее состав входит блок рецепторов, выделяющих определенную категорию вход-50

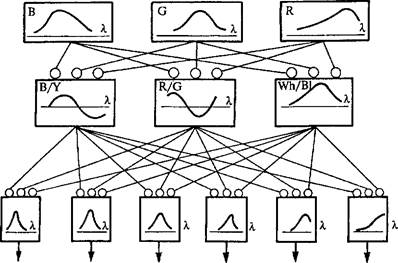
Рис. 12. Структура анализатора цвета-
В, О, Я (сине-, зелено- и красночувствительные колбочки) — I уровень обработки. Предетекторы представляют II уровень обработки сигнала в анализаторе цвета. III уровень — карта селективных к различным цветам детекторов, имеющих разные синаптические связи с предетекторами; кривые в квадратах — характерные ответы нейронов при изменении стимула; \ — длина волны (по С.В. Фомину и соавт., 1979).
ных сигналов (например, излучение определенной длины волны:
В, С, К). Вторым блоком является блок предетекторов. Они трансформируют сигналы рецепторов в форму, пригодную для селективного возбуждения детектора. Для нейронной системы, обеспечивающей цветовосприятие, функцию предетекторов выполняет ансамбль нейронов, представленный двумя типами цветооппо-нентных клеток (К/С и В/У), а также двумя типами ахроматических нейронов: яркости и темноты (\УЬ/В1). Предетекторы си-наптически связаны со всеми нейронами-детекторами цвета параллельно (рис. 12).
При действии стимула у элементов ансамбля нейронов в силу их разной чувствительности возникают разные по силе и комбинации возбуждения. Комбинация возбуждений в ансамбле нейронов получила название «вектор возбуждения». Ансамбль нейронов, конвергирующих на нейроны-детекторы, реагируя на стимул возбуждением, проецирует свой вектор возбуждения на все нейроны-детекторы, выделяющие соответствующее качество стимула. С уровня
предетекторов на цветовые детекторы поступает четырехкомпонентный вектор возбуждения от 4 типов предетекторов (нейронов К/О, В/У, ^Ь,В1). Синапсы, через которые предетекторы связываются с разными цветовыми детекторами, различаются по своей эффективности. Сила синоптической связи каждого детектора цвета с предетекторами характеризует его индивидуальный вектор си-наптических связей (или вектор связи). Реакция детектора на стимул зависит от вектора его синаптических связей с предетекторами. Поэтому Е.Н. Соколов предлагает рассматривать возбуждение детектора как результат суммации попарных произведений, образованных возбуждениями предетекторов и эффективностью воздействия (весами) соответствующих им синапсов на данном детекторе. Сумма таких попарных произведений образует скалярное произведение двух векторов', вектора возбуждения (возбуждения предетекторов) и вектора связи, представленного весами синапсов на данном детекторе.
Вектор возбуждения, возникающий в ансамбле нейронов, конвергирующих на детектор, характеризуется силой возбуждения (длиной вектора) и его направленностью в виде соотношения возбуждений внутри ансамбля нейронов (направлением вектора).
Предполагается, что существует операция нормирования вектора возбуждения, в результате чего все стимулы, воздействующие на нейронный ансамбль, кодируются равными по длине векторами возбуждения. Эта операция осуществляется за счет следующих процедур: возбуждения элементов ансамбля суммируются; с ростом уровня суммарного возбуждения снижается реакция элементов ансамбля; существует элемент, создающий шум, который исчезает с появлением сигнала, при этом величина подавления пропорциональна интенсивности стимула.
Вектор связи у каждого детектора — величина постоянная, так как синапсы на них от предетекторов относятся к непластичным.
Таким образом, скалярное произведение вектора возбуждения предетекторов и вектора их связи с детектором зависит лишь от ориентации этих векторов. При их совпадении нейрон-детектор реагирует на стимул максимально. И на карте детекторов возникает рельеф возбуждения с максимумом на том детекторе, вектор связи которого совпадает по направлению с возникшим вектором возбуждения. С изменением стимула вектор возбуждения меняется и максимум рельефа возбуждения смещается на другой детектор. Локус максимального возбуждения на карте детекторов кодирует стимул.
Каким образом признаки стимула, выделяемые отдельными группами нейронов-детекторов, интегрируются в целостный образ воспринимаемого объекта? Несмотря на большие успехи, дос-52
;:;^, тигнутые при изучении нейронных процессов и функций различных групп нейронов, наука располагает значительно меньшими данными об интегративных механизмах мозга. В этой связи нейро-биология все настоятельнее обращается к проблеме целостности и тому, как она была представлена в системе гештальтпсихологии. Термин «гештальт» был введен, чтобы подчеркнуть качественное своеобразие восприятия паттерна элементов, которое не сводится к их сумме. Примером гештальта может служить открытие кажущегося движения (фи-феномена). При последовательной подаче двух точек, пространственно разделенных, человек видит движение одной точки по линии, соединяющей их, — явление, не сводимое к восприятию последовательности двух пространственно разделенных точек.
С позиции теории векторного кодирования информации появлению целостного образа, гештальта, соответствует определенный вектор. Он качественно отличен от других векторов специфическим соотношением своих компонентов (Соколов Е.Н., 1996). Компонентами такого вектора могут быть возбуждения нейронов-детекторов отдельных признаков, которые конвергируют на гностической единице — нейроне более высокого порядка, избирательно. реагирующем на сложные изображения (лица, жесты и т.д.). Таким образом, формирование гештальта связано с участием гностических единиц. При этом объединение детекторов элементарных признаков происходит не в результате установления между ними горизонтальных связей (одноуровневых), а вследствие включения их в иерархически организованную нейронную сеть. Возникновение целостного субъективного образа предмета связано с пирамидой последовательных нейронных преобразований, вершиной которой является операция, выполняемая гностической единицей (Соколов Е.Н., 1996). Отдельным зрительным гештальтам соответствуют отдельные гностические единицы.
В организации гештальтов восприятия существенную роль выполняет память. При рассмотрении объекта глаза последовательно фиксируют наиболее информативные точки. Цепочка таких фиксаций интегрируется в гештальт на основе иконической памяти. Восприятие музыки (мелодии, аккордов и т.д.) также связано с формированием гештальтов. В этом процессе участвует кратковременная память. В ней некоторое время удерживаются не только комплексы элементов, но и их временные параметры: длительность звучания элементов, интервалы между ними. Звуковой гештальт — целостное образование, хотя оно и имеет свою протяженность во времени. В связи с этим появился новый термин — «звуковые объекты».

«Сенситивныйо период
Рис. 13. Схематическое изображение роли «сенситивпого» периода в формировании долговременных пластических перестроек синапсов на гностической единице О. под влиянием активации.
А — активация; 0^, Сг, 0^,, 0^ — гностические единицы; 5. 5,, 5. — стимулы, достигающие гностических единиц (по Е.Н. Соколову, 1996).
Сами гностические единицы являются продуктом обучения. По данным некоторых авторов, у обезьян их находят в передней части вентральной височной коры. В опытах Я. Мияшиты (М1уа5Ька V. е1 а1., 1991) обезьян обучали узнавать сходство тестового стимула с эталоном, хранящимся в памяти. Для этого обезьяне предъявляли 97 фигур, различающихся формой, цветом, величиной, ориентацией. Стимулы генерировались компьютером и предъявлялись на мониторе. Животному сначала показывали образец, затем через 16 с давали тестовый стимул. Если обезьяна обнаруживала их сходство и об этом сообщала двигательной реакцией, то получала подкрепление. После обучения в передней вентральной височной коре были найдены нейроны, избирательно реагирующие на стимулы, которые использовались в опыте. Применение нового набора из 97 стимулов, генерируемых компьютером, не обнаружило каких- либо нейронов с селективной настройкой на их восприятие, т.е. в процессе обучения — выработки инструментального рефлекса — для каждого из 97 стимулов сформировалась своя гностическая единица, избирательно на него реагирующая. Процесс формирования гностических единиц напоминает изменение свойств у нейронов-детекторов в сенситивный период.
Модель последовательного формирования избирательно реагирующих гностических единиц Е.Н. Соколов описывает следующим образом. В передней вентральной височной коре имеется пул
резервных нейронов, которые слабо реагируют на разные стимулы. Под влиянием сигнала новизны из гиппокампа они активируются, включая на короткое время механизм пластических перестроек в синапсах. Это и создает у нейронов «сенситивный период» (рис. 13). В этих условиях сигнал, поступающий от детекторов к резервному нейрону, повышает синаптическую связь между ними. В результате нейрон становится селективно настроенным на восприятие данного стимула. Селективный период обрывается с завершением процесса обучения, и синаптические связи нейрона перестают меняться. Появление нового стимула активирует очередной резервный нейрон и т.д. Таким образом, анализ отдельных зрительных характеристик стимула выполняется нейронами-детекторами. Выделенные элементарные признаки синтезируются в долговременной памяти в передневентральной височной коре на гностических нейронах. Выполняется принцип «один гештальт — один нейрон». Отображение стимула в гностической единице характеризуется высокой степенью абстракции. При изменении размера, ориентации, цвета объекта гностический нейрон продолжает сохранять избирательность своей реакции на данный объект. Осознание объекта в процессе его восприятия связывают с дополнительной активацией гештальт-пирамиды, через механизм синхронизации электрической активности мозга в диапазоне гамма-частот (см. главу «Сознание»).
Глава 5 ВНИМАНИЕ
|
|
|
