 |


Происхождение человеческих рас.
|
|
|
|
Расы человека, по-видимому, появились сравнительно недавно. Согласно одной из схем, основанной на данных молекулярной биологии, разделение на два больших расовых ствола - негроидный и европеоидно-монголоидный - произошло скорее всего около 100 тысяч лет назад, а дифференциация европеоидов и монголоидов - около 45-60 тысяч лет назад. Большие расы в основном формировались под влиянием природных и социально-экономических условий в ходе внутривидовой дифференциации уже сложившегося человека разумного, начиная с эпохи позднего палеолита и мезолита, но главным образом - в неолите. Европеоидный тип установился с неолита, хотя отдельные его черты прослеживаются в позднем или даже среднем палеолите. Фактически отсутствуют достоверные свидетельства присутствия монголоидов в Восточной Азии в донеолитическую эпоху, хотя в Северной Азии они, возможно, существовали уже в позднем палеолите. В Америке предки индейцев не были сложившимися монголоидами. Также и Австралия заселялась еще «нейтральными» в расовом отношении неоантропами.
Расы и этнос.
Большие расы человека занимают обширные территории, охватывающие народы, которые различаются по уровню экономического развития, культуре, языку. Нет сколько-нибудь четких совпадений понятий «раса» и «этнос» (народ, нация, национальность). В то же время есть примеры антропологических типов (малые, а иногда и большие расы), которые соответствуют одному или нескольким близким этносам, например, лапаноидная раса и саамы. Гораздо чаще, однако, наблюдается обратное: один антропологический тип широко распространен среди многих этносов, как, например, в коренном населении Америки или у народов Северной Европы. Вообще все крупные народы, как правило, неоднородны в антропологическом отношении. Нет также совпадения между расами и языковыми группами - последние возникали позже, чем расы. Так, среди тюркоязычных народов имеются представители как европеоидов (азербайджанцы), так и монголоидов (якуты). Термин «расы» неприменим к языковым семьям - например, нужно говорить не о «славянской расе», а о группе родственных народов, говорящих на славянских языках.
|
|
|
Расы и расизм. Многие расовые признаки имеют приспособительное значение. Например, у представителей экваториальной расы темная пигментация кожи защищает от обжигающего действия ультрафиолетовых лучей, а вытянутые пропорции тела увеличивают отношение поверхности тела к его объему и тем самым облегчают терморегуляцию в условиях жаркого климата. Однако расовые признаки не являются определяющими для существования человека, поэтому они ни в коей мере не свидетельствуют о каком-либо биологическом или интеллектуальном превосходстве или, напротив, неполноценности той или иной расы. Все расы находятся на одном и том же уровне эволюционного развития и характеризуются одинаковыми видовыми особенностями. Поэтому концепции о якобы неравноценности человеческих рас в физическом и психическом отношениях (расизм), выдвигаемые с середины 19 века, научно несостоятельны. Расизм имеет отчетливые социальные корни и всегда использовался как оправдание для насильственного захвата земель и дискриминации коренных народов. Расисты обычно игнорируют тот факт, что различия между достижениями разных народов полностью объясняются историей их культур, зависящей от внешних факторов, от их исторически меняющейся роли. Достаточно сравнить уровень культурного развития населения Северной Европы в наши дни и в эпоху великих цивилизаций прошлого в Месопотамии, Египте, долине Инда.
|
|
|
Человечество это один вид, спору нет. Но внутри него есть разновидности, отрицать существование которых - это... совершенно правильная политика многонациональных государств.

Лекция 14. Филогенез.
Закон Бэр а - закон зародышевого сходства: «Эмбрионы последовательно переходят в своем развитии от общих признаков типа ко все более специальным признакам. Позднее всего развиваются признаки, указывающие на принадлежность эмбриона к определенному роду, виду, и, наконец, развитие завершается появлением характерных особенностей данной особи». Бэр не придавал этому «закону» эволюционного смысла (он до конца жизни так и не принял эволюционного учения Дарвина), однако позднее этот закон стал рассматриваться как «эмбриологическое доказательство эволюции» и свидетельство происхождения животных одного типа от общего предка.
Биогенетический закон Геккеля - Мюллера (также известен под названиями «закон Геккеля», «закон Мюллера - Геккеля», «закон Дарвина - Мюллера - Геккеля», «основной биогенетический закон»): каждое живое существо в своем индивидуальном развитии (онтогенез) повторяет в известной степени формы, пройденные его предками или его видом (филогенез).

Рисунок из книги Ремане (1892), воспроизводящий исходную иллюстрацию Геккеля.
В настоящее время в своем исходном виде не признается современной биологической наукой. По современной трактовке биогенетического закона, предложенной русским биологом А. Н. Северцовым в начале 20 века, в онтогенезе происходит повторение признаков не взрослых особей предков, а их зародышей.
Опираясь только на основной биогенетический закон, невозможно объяснить процесс эволюции: бесконечное повторение пройденного само по себе не рождает нового. Так как жизнь существует на Земле благодаря смене поколений конкретных организмов, эволюция ее протекает благодаря изменениям, происходящим в их онтогенезах. Эти изменения сводятся к тому, что конкретные онтогенезы отклоняются от пути, проложенного предковыми формами, и приобретают новые черты.
К таким отклонениям относятся, например, ценогенезы - приспособления, возникающие у зародышей или личинок и адаптирующие их к особенностям среды обитания. У взрослых организмов ценогенезы не сохраняются. Примерами ценогенезов являются роговые образования во рту личинок бесхвостых земноводных, облегчающие им питание растительной пищей. В процессе метаморфоза у лягушонка они исчезают и пищеварительная система перестраивается для питания насекомыми и червями. К ценогенезам у амниот относят зародышевые оболочки, желточный мешок и аллантоис, а у плацентарных млекопитающих и человека - еще и плаценту с пуповиной.
|
|
|
Ценогенезы, проявляясь только на ранних стадиях онтогенеза, не изменяют типа организации взрослого организма, но обеспечивают более высокую вероятность выживания потомства. Они могут сопровождаться при этом уменьшением плодовитости и удлинением зародышевого или личиночного периода, благодаря чему организм в постэмбриональном или постличиночном периоде развития оказывается более зрелым и активным. Возникнув и оказавшись полезными, ценогенезы будут воспроизводиться в последующих поколениях. Так, амнион, появившийся впервые у предков пресмыкающихся в каменноугольном периоде палеозойской эры, воспроизводится у всех позвоночных, развивающихся на суше, как у яйцекладущих - пресмыкающихся и птиц, так и у плацентарных млекопитающих.
Другой тип филогенетически значимых преобразований филогенеза - филэмбриогенезы. Они представляют собой отклонения от онтогенеза, характерного для предков, проявляющиеся в эмбриогенезе, но имеющие адаптивное значение у взрослых форм. Так, закладки волосяного покрова появляются у млекопитающих на очень ранних стадиях эмбрионального развития, но сам волосяной покров имеет значение только у взрослых организмов.
Такие изменения онтогенеза, будучи полезными, закрепляются естественным отбором и воспроизводятся в последующих поколениях. В основе этих изменений лежат те же механизмы, которые обусловливают врожденные пороки развития: нарушение пролиферации клеток, их перемещения, адгезии, гибели или дифференцировки. Однако от пороков их так же, как и ценогенезы, отличает адаптивная ценность, т.е. полезность и закрепленность естественным отбором в филогенезе.
|
|
|
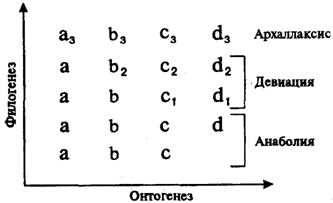
В зависимости от того, на каких этапах эмбриогенеза и морфогенеза конкретных структур возникают изменения развития, имеющие значение филэмбриогенезов, различают три их типа.
1. Анаболии, или надставки, возникают после того, как орган практически завершил свое развитие, и выражаются в добавлении дополнительных стадий, изменяющих конечный результат.
К анаболиям относят такие явления, как приобретение специфической формы тела камбалой лишь после того, как из икринки вылупляется малек, неотличимый от других рыб, а также появление изгибов позвоночника, сращение швов в мозговом черепе, окончательное перераспределение кровеносных сосудов в организме млекопитающих и человека.
2. Девиации - уклонения, возникающие в процессе морфогенеза органа. Примером может являться развитие сердца в онтогенезе млекопитающих, у которых оно рекапитулирует стадию трубки, двухкамерное и трехкамерное строение, но стадия формирования неполной перегородки, характерной для пресмыкающихся, вытесняется развитием перегородки, построенной и расположенной иначе и характерной только для млекопитающих. В развитии легких у млекопитающих также обнаруживается рекапитуляция ранних стадий предков, позднее морфогенез идет по - новому.
|
|
|
