 |


Влияние окружающей среды на поведение
|
|
|
|
Развитие поведения
Типы поведения животных складываются в результате сложного взаимодействия внешних стимулов и состояния внутренней среды организма. Однако любой тип поведения определяется тем, каким образом животное обрабатывает поступающую к нему информацию. Внутренние системы обработки информации формируются в процессе развития от оплодотворенного яйца к зародышу и взрослому животному; этот процесс называется онтогенезом. Изучая онтогенез, мы можем выявить способы взаимодействия генетической информации и информации, полученной из окружающей среды (или путем научения), что в конечном счете формирует соответствующий тип поведения животных.
Онтогенез
Почти все живые организмы построены из одних и тех же веществ, причем различия между видами определяются различиями в способах объединения этих основных веществ. Механизмы, контролирующие такое объединение, изучены слабо, однако известно, что гены контролируют развитие, детерминируя синтез белков, которые регулируют сложную организацию процессов онтогенеза.
Однако эти регуляторные белки эффективны лишь настолько, насколько развивающийся организм чувствителен к информации, которую они несут. Действие регуляторной молекулы основано не на создании некой системы, соответствующей какому-то определенному плану, напротив, эта молекула вызывает специфическую реакцию в клетках, на поверхности которых находятся рецепторы для регуляторных молекул. Таким образом, ответ развивающегося зародыша на продукт регуляторного гена зависит от внутренней организации зародыша ко времени образования этого регулятора. Кроме того, этот регулятор функционирует только при определенных условиях внутренней среды организма. В качестве примера рассмотрим сначала некоторые аспекты эмбриогенеза у видов с относительно простым поведенческим репертуаром.
|
|
|
Aplysia - лишенная раковины морская улитка, масса которой может достигать 4,5 кг. Как типичная улитка, Aplysia - настоящийгермафродит. Оплодотворение у нее внутреннее, и обычно образуются цепи, включающие до 10 и более копулирующих животных, причем каждая улитка оплодотворяет соседнюю, в то время как сама оплодотворяется другой. После оплодотворения Aplysia откладывает более миллиона яиц, соединенных в длинный шнур. Сначала шнур выталкивается из репродуктивного мешка сокращением мышц последнего. Когда шнур достигает достаточной длины, улитка захватывает его конец в складку верхней губы и, двигая головой взад и вперед, помогает высвобождению яиц из репродуктивного мешка (рис. 3.1). Специальная железа секретирует клейкое вещество, с помощью которого шнур склеивается, образуя более компактную массу в виде беспорядочного мотка. Наконец, улитка крепко прижимает головой эту массу яиц к твердому субстрату (рис. 3.1), где они и остаются до вылупления. Вся эта серия последовательных действий при откладке яиц-жест-ко зафиксированный поведенческий стереотип. Во время этого процесса у животного увеличивается частота сокращений

|
| Рис. 3.1. Поведение Aplisvu, откладывающей яйца. Лента яиц в защитной оболочке выталкивается из полового протока, выходное отверстие которого расположено на боку тела. Животное захватывает этот шнур в складку верхней губы (слева) и прикрепляет его к субстрату (справа). (По Scheller, Axel, 1981.) |
сердца и частота дыхательных движений, оно полностью прекращает питаться и передвигаться.
Шеллер и Аксель (Scheller, Axel, 1981) обнаружили, что эта последовательность действий контролируется генами на разных уровнях, как во время эмбриогенеза, так и в процессе ее осуществления. Клетки, которые дают начало всей нервной системе аплизии, являются потомками нескольких клеток, располагающихся в стенке тела зародыша. На ранних стадиях развития эти преднервные клетки синтезируют специфический белок, названный гормоном откладывания яиц (ELH-egg-laying hormone). Полагают, что все эти преднервные клетки являются потомками единственной клетки зародыша, в которой реализуется способность соответствующего гена кодировать синтез ELH. Во всех других клетках зародыша также имеется этот ген, но в них он не функционирует. Клетки, синтезирующие ELH, продолжают делиться и перемещаются к своему окончательному местоположению в развивающемся организме, где дифференцируются в нервные клетки взрослой особи. Поэтому у взрослой улитки некоторые нервные клетки наследуют способность синтезировать ELH, а другие - нет.
|
|
|
Ко времени полового созревания все эти системы улитки занимают соответствующее им местоположение. В надлежащее время начинают вырабатываться другие гормоны, которые и приводят клетки, синтезирующие ELH, в состояние полной работоспособности. Выделяющийся ELH заставляет возбуждаться соседние нейроны (т. е. в данном случае гормон действует как нейромедиатор). Этот гормон циркулирует и в крови, заставляя сокращаться специфические мышечные волокна и действуя, таким образом, как гормон; в результате приводится в действие сложный комплекс координированного поведения откладки яиц. На примере таких фиксированных действий отчетливо видна вся важность координации процессов эмбриогенеза, активности генов, действия нейромедиаторов, нервов и мышц, приводящей к реализации поведения, столь существенного для размножения улитки.
У аплизии гены непосредственно контролируют развитие стереотипных форм поведения. И если гены на каком-то уровне (например, в эмбриогенезе, когда происходит закладка общего плана строения) контролируют становление поведения в целом, то различные средовые факторы могут в разной степени модифицировать те или иные формы поведения, появившиеся в эмбриогенезе. Фенотипический эффект одних и тех же генов может быть различен в зависимости от того, в каких условиях развивается животное. Например, у ракообразного Gammarus глаза обычно красного цвета. Форд и Хаксли (Ford, Huxley, 1927) обнаружили у Gammarus мутацию, которая влияет на скорость образования глазного пигмента во время определенной стадии онтогенеза. Если на этой стадии развития температуру
|
|
|
окружающей среды повысить до некоторого уровня, то глаза остаются красными, при более высоких температурах они становятся шоколадно-коричневыми, а при промежуточных температурах - промежуточного цвета. Учитывая огромную сложность биохимических процессов, принимающих участие в экспрессии генов, в процессах роста и дифференцировки клеток (Ham, Veomett, 1980), не приходится удивляться, что среда, в которой протекают данные процессы, влияет на развитие этих клеток. Присущие животным стабилизирующие, или регуляторные механизмы в какой-то степени исправляют отклонения и контролируют скорость и направление процессов развития. Иногда одна и та же структура или один и тот же тип поведения развиваются разными путями - явление, известное под названием эквифинальностъ. Тем не менее отклонения от обычного пути развития неизбежны. В некоторых случаях эти отклонения могут быть летальными, в других же они приводят к возникновению различий между особями, несущими один и тот же генотип.
Некоторые ранние этологи считали, что гены непосредственно детерминируют поведение. Так, Лоренц (Lorenz, 1965) полагал, что генетическое детерминирование аналогично архитектурному проекту здания. Генетический проект представляет собой некий план создания взрослого животного. Признавая, что для строительства здания нужны кирпичи, раствор и рабочая сила, Лоренц определил четкие различия между условиями, необходимыми для воплощения проекта в здание, и информацией, которая содержится в проекте и от которой зависят характеристики готового сооружения. Исследователи, критиковавшие эту точку зрения (например, Lehrman, 1970), считали, что если архитектурный проект изоморфен строению, которое он представляет, то этого нельзя сказать в случае генетического проекта. Ведь между измерениями, указанными на архитектурном проекте, и измерениями построенного здания существует однозначное соответствие, какого не может быть в онтогенезе. Даже если верно то, что в каждом гене закодирован определенный ключевой фермент, все же это сильно отличается от проекта. Представление о том, что информацию, получаемую в процессе развития от генов, можно отделить от информации, получаемой из окружающей среды, было критически воспринято Хеббом (Hebb, 1953), который считал, что спрашивать, насколько данный тип поведения зависит от генетических, а насколько от средовых факторов, - это все равно, что спрашивать, насколько площадь поля зависит от его длины, а насколько - от ширины.
|
|
|
Большой интерес исследователей к изучению онтогенеза поведения объяснялся их желанием продемонстрировать разнообразие путей, по которым развиваются различные типы поведения. Оказалось, что существует целый спектр процессов, различающихся по влиянию на них факторов окружающей среды, - от относительно независимых до полностью зависимых.
Генетический детерминизм ранних этологов привел к признанию того, что влияние генов и влияние окружающей среды сложным образом связано друг с другом, однако связь эта не такая, как полагал Лоренц (Lorenz, 1965), - связь проекта и построенного на его основе здания; скорее она носит эпигенетический характер, т.е. каждая стадия развития создает основу для следующей стадии, а не предопределяет ее. По мнению Брауна (Brown, 1975), процессы эпигенеза суммарно можно представить следующим образом. Начиная с оплодотворенного яйца, или зиготы (P1), фенотип организма во время следующей стадии развития (Р2) будет определяться генами, которые детерминируют его рост и дифференцировку в данный период (G1), и условиями среды (E1), в которой протекает развитие; т. е. зигота плюс гены плюс окружающая среда будут определять фенотип на следующей стадии:

Фенотип (Р3) на следующей стадии развития будет определяться взаимодействием фенотипа Р2, генов G2 и условий окружающей среды (Е2); это приведет к развитию фенотипа Р3:
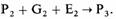
Эта запись показывает, что все три элемента - ген, окружающая среда и исходный фенотип - играют определенную роль в нормальном развитии. Развитие может достичь следующей стадии только в том случае, если развивающееся животное обладает соответствующим фенотипом, если у него синтезируются правильные генные продукты и если условия среды не выходят за рамки определенных границ.
На ранних стадиях развития (P1 + G1 + Е1 --> Р2) средовой компонент E1 представляет собой в основном биохимическое окружение, в котором находится ранний зародыш. На более поздней стадии развития (Р2 + G2 + Е2 --> Р3) средовой компонент Е2 может быть представлен веществами яйца. После вылупления из яйца или рождения окружающая среда животного включает информацию, получаемую его органами чувств, а также биохимические вещества, получаемые в процессе питания и другими способами. В этой ситуации может иметь место научение, которое постоянно влияет на поведенческий фенотип (Р3) животного.
|
|
|
Связь между врожденным и приобретенным поведением удобно изучать, сравнивая птенцов выводковых (у которых вылупление происходит на поздних стадиях развития) и птенцовых (у которых вылупление имеет место на ранних стадиях) видов птиц. Если сравнивать птенцов этих птиц на одной и той же стадии развития, например на стадии, когда появляются первые перья, то оказывается, что птенец выводковой птицы находится в это время еще в яйце, тогда как птенец птенцовой птицы уже вылупился. Ко времени вылупления выводковой птицы молодая птенцовая имела возможность уже многому научиться, тогда как выводковый птенец учиться еще и не начинал. И что особенно интересно - мы обнаружили, что выводковая птица может обладать такими врожденными навыками которые птенцовой птице приходится осваивать. Например, белоголовая воробьиная овсянка, относящаяся к птенцовым птицам, заучивает видоспецифическую песню с первых дней своей жизни в гнезде. Напротив, птицы, относящиеся к выводковым, проходят ранние стадии развития в яйце и при вылуплении уже обладают способностью к нормальной вокализации, хотя раньше никогда не слышали звуки, издаваемые родителями.
Мы склонны думать, что происходящие внутри яйца события представляют собой лишь физические процессы созревания; иными словами, мы считаем, что факторы, влияющие на этот процесс, предопределены. Однако мы не должны забывать, что нормальная среда белоголовой воробьиной овсянки также предопределена в том смысле, что в критический для научения песне период птенцы в гнезде обычно слышат песню своего вида.
На последующих стадиях развития Р3 + G3 + Е3 --> Р4. К этой стадии влияние приобретенного опыта будет заметно как у птенцовых, так и у выводковых птиц. Поведение, которое несомненно складывается под влиянием опыта, нельзя назвать «врожденным». Однако здесь есть небольшое принципиальное отличие от ситуации, складывающейся на ранних стадиях развития. Степень предопределенности средовых факторов зависит в основном от изменчивости окружающей среды. Внутри яйца среда обычно изменяется незначительно, поэтому мы склонны считать поведение только что вылупившегося птенца врожденным. Однако вне яйца среда также может изменяться очень незначительно, и это зависит от экологических условий, типичных для вида. Таким образом, постнатальное поведение птенцовых птиц может быть точно так же предопределено, как и постнатальное поведение выводковых. Если с эволюционной точки зрения важно, чтобы развитие давало какой-то конкретный конечный результат, как это отчетливо видно в случае с песней у воробьиных птиц, то мы можем ожидать, что влияние окружающей среды будет таким, чтобы обеспечить этот результат.
Влияние окружающей среды на поведение
Некоторые формы поведения проявляются только на определенных стадиях развития. Иногда такое поведение развивается, по-видимому, без какой бы то ни

|
| Рис. 4.7. Самка полевого сверчка на Y-образном лабиринте. Ее движения по лабиринту показывают, призывному сигналу какого самца она оказывает предпочтение. (По Ноу, 1974.) |
было заметной тренировки. Например, голуби в определенном возрасте начинают махать крыльями и неуверенно летать, но с практикой их летательные способности улучшаются. Однако Громан (Grohmann, 1939) в ставшем теперь классическим эксперименте выращивал группу голубей в трубах, в которых они не могли двигать крыльями. Другая группа такого же возраста развивалась без ограничений. Когда эти развивавшиеся на свободе голуби достигли стадии, на которой они могли удовлетворительно летать, были освобождены голуби из труб. Оказалось, что они также способны летать сразу же после освобождения. У мутантных цыплят, лишенных оперения, развиваются нормальные вестибулярные рефлексы, проявляющиеся в движениях крыльев, хотя эти рефлексы совершенно бесполезны (Provine, 1981). На первый взгляд кажется очевидным, что поведение не зависит от средовых факторов, но не следует считать, что способность к полету развивается независимо от онтогенетических возможностей. Вероятнее все же, способность к полету зависит от некоторых существенных условий, хотя наши знания о них фрагментарны (Ham, Veomett, 1980). Сходный пример - вокализация у голубей и домашних кур, которая весьма стереотипна и проявляется на определенных стадиях развития. Она зависит не от слухового опыта, а от определенного гормонального состояния, являющегося частью общего созревания.
Многие типы движений развиваются при очевидном отсутствии тренировок или примера (Hinde, 1970). Однако результаты изучения некоторых форм поведения в эмбриогенезе позволили предположить, что в них фрагментарно и неполно включены движения, которые, возможно, влияют на процесс развития (например, Кио, 1932, 1967; Kraijt, 1964; Anthoney, 1968). В качестве примера мы рассмотрим работу Куо (Кио, 1932) по развитию поведения у куриных зародышей. Куо делал окна в скорлупе яйца и непосредственно наблюдал за поведением зародыша (рис. 3.2). Он обнаружил, что зародыш постоянно подвергается воздействию различных сигналов, возникающих как внутри яйца в результате его собственной активности, так и поступающих снаружи. Можно было наблюдать различные движения, в том числе начальные пассивные движения головы, вызванные сокращениями сердца. Активные движения головы развиваются в течение нескольких дней и сопровождаются открыванием и закрыванием клюва. К семнадцатому дню наблюдаются движения, похожие на клевание. Некоторые исследователи полагают, что таким образом цыпленок учится клевать (например, Lehrman, 1953; Schneirla, 1965). Другие (например, Thorpe, 1963; Lorenz, 1965) поднимают эту идею на смех. Независимо от того, имеет место научение или нет, не приходится сомневаться в том, что развивающийся зародыш отвечает на различные сигналы, поступающие как изнутри яйца, так и из внешней среды, и что эти сигналы влияют на эмбриональное развитие. Так, Маргарет Винс (Margaret Vince, 1969) показала, что сигналы, исходящие от зародыша, могут влиять на развитие других яиц в гнезде, синхронизируя вылупление. Готтлиб (Gottlieb, 1971) обнаружил, что утиные зародыши начинают реагировать на сигналы матери за пять дней до вылупления и что такой опыт может влиять на последующее поведение утят. Однако другие исследователи (например, Hamburger, 1963) получили данные, свидетельствующие о том, что движения у куриных за-

|
| Рис. 3.3. Утенок, следующий за своей "матерью» в аппарате, сконструированном для изучения различных аспектов запечатления. (По Gottlieb, 1971.) |
родышей развиваются при отсутствии сенсорной стимуляции, и что на типы движений, которые проявляются позднее в онтогенезе, не обязательно оказывают влияние движения раннего зародыша (см. Gottlieb, 1970).
Читатели, должно быть, помнят, что потомство разных видов развивается разными путями. Если утята и козлята способны бегать сразу же после появления на свет, то птенцы черного дрозда и котята в этом возрасте беспомощны. Нас не удивляет утверждение о том, что на постна-тальное развитие последних влияет опыт. Так должны ли мы удивляться тому, что животное, находящееся в яйце или в утробе матери и проходящее такие же стадии общего развития, подвергается воздействию определенных сигналов. По-видимому, нет принципиальной разницы между защитными приспособлениями, окружающими утенка в яйце, и защитой молодых черных дроздов в гнезде. Точно так же, по-видимому, невелики принципиальные различия между влиянием среды в том и в другом случае. Для нормального развития потомства каждого вида важны определенные типы жизненного опыта. Например, взрослая кошка не будет обладать нормальным зрением, если в течение первых месяцев жизни у нее возникают какие-либо нарушения в координации зрения двумя глазами, например, при искусственном косоглазии или при выключении каждого глаза поочередно (через день), так чтобы оба глаза не функционировали вместе (Hubel, Wiesel, 1965). Опыт, который необходим для нормального развития животного, различен у разных видов.
|
|
|
