 |


36. Ввнутриклеточный обмен липидов (катаболизм триацетилглицеролов, окисление жирных кислот)
|
|
|
|
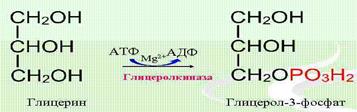 Катаболизм триацилглицеролов. Липолиз (гидролиз) резервных липидов в периферических тканях катализируется гормончувствительной липазой до глицерола и свободных высших жирных кислот. Наиболее активно этот процесс идет в жировой ткани, которая распространена по всему организму: под кожей, в брюшной полости, образует жировые прослойки вокруг отдельных органов. Свободные жирные кислоты либо вновь вовлекаются в синтез липидов, либо подвергаются р-окислению, либо диффундируют в плазму крови, где связываются с сывороточным альбумином и транспортируются в другие ткани, являясь одним из основных источников энергии. Глицерол в жировой ткани практически не утилизируется. Он диффундирует в плазму крови, оттуда поступает в такие ткани, как печень или почки, где фосфорилируется под действием активной глицеролкиназы при участии АТФ. Глицерол-З-фосфат (активированная форма глицерола) дегидрируется, при действии НАД+-зависимой глицеролфосфатдегидрогеназы, а образующиеся триозофосфаты либо далее метаболизируют по пути гликолиза, либо вовлекаются в процесс глюконеогенеза (синтез глюкозы):
Катаболизм триацилглицеролов. Липолиз (гидролиз) резервных липидов в периферических тканях катализируется гормончувствительной липазой до глицерола и свободных высших жирных кислот. Наиболее активно этот процесс идет в жировой ткани, которая распространена по всему организму: под кожей, в брюшной полости, образует жировые прослойки вокруг отдельных органов. Свободные жирные кислоты либо вновь вовлекаются в синтез липидов, либо подвергаются р-окислению, либо диффундируют в плазму крови, где связываются с сывороточным альбумином и транспортируются в другие ткани, являясь одним из основных источников энергии. Глицерол в жировой ткани практически не утилизируется. Он диффундирует в плазму крови, оттуда поступает в такие ткани, как печень или почки, где фосфорилируется под действием активной глицеролкиназы при участии АТФ. Глицерол-З-фосфат (активированная форма глицерола) дегидрируется, при действии НАД+-зависимой глицеролфосфатдегидрогеназы, а образующиеся триозофосфаты либо далее метаболизируют по пути гликолиза, либо вовлекаются в процесс глюконеогенеза (синтез глюкозы):
 Гормоночувствительная липаза является важнейшим регуляторным ферментом процессов липолиза. Многие гормоны являются активаторами этого фермента. К гормонам, которые быстро промотируют липолиз, относятся прежде всего катехоламины (адреналин и норадреналин) и глюкагон, которые стимулируют активность аденилатциклазы — фермента, катализирующего образование из АТФ циклического АМФ (цАМФ).
Гормоночувствительная липаза является важнейшим регуляторным ферментом процессов липолиза. Многие гормоны являются активаторами этого фермента. К гормонам, которые быстро промотируют липолиз, относятся прежде всего катехоламины (адреналин и норадреналин) и глюкагон, которые стимулируют активность аденилатциклазы — фермента, катализирующего образование из АТФ циклического АМФ (цАМФ).
Окисление жирных кислот. Окислительное расщепление жирных кислот — универсальный биохимический процесс, протекающий во всех видах живых организмов. У млекопитающих этот процесс происходит во многих тканях, в первую очередь в печени, почках, сердечной и скелетной мышцах. В клетке окисление жирных кислот локализовано в матриксе митохондрий. В настоящее время биохимические превращения жирных кислот в процессе р-окисления детально изучены. Они включают следующие основные этапы:
|
|
|
• активацию жирной кислоты в цитоплазме клетки. Активация жирной кислоты является двухстадийным процессом.
Первая стадия: RH2СН2СООН + АТФ → RCН2СН2СO~АМФ + H5P2O7
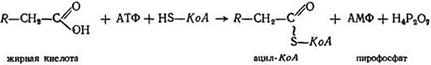 Вторая стадия: RСН2СН2CO~SКоА + HS~КоA= RСН2СН2C-O~SКоА + АМФ. Суммарная реакция:
Вторая стадия: RСН2СН2CO~SКоА + HS~КоA= RСН2СН2C-O~SКоА + АМФ. Суммарная реакция:
 При действии фермента пирофосфатазы в пирофосфате расщепляется богатая энергией фосфоангидридная связь, что обеспечивает полноту протекания процесса активации и делает эту реакцию необратимой: Н4Р2О7+ Н2О =(фермент пирофосфатаза) 2Н3РО4 Таким образом, для активации одной молекулы жирной кислоты расходуются две макроэргические связи АТФ.
При действии фермента пирофосфатазы в пирофосфате расщепляется богатая энергией фосфоангидридная связь, что обеспечивает полноту протекания процесса активации и делает эту реакцию необратимой: Н4Р2О7+ Н2О =(фермент пирофосфатаза) 2Н3РО4 Таким образом, для активации одной молекулы жирной кислоты расходуются две макроэргические связи АТФ.
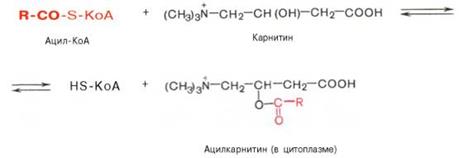
• транспорт ацильной группы в митохондрии: Внутренняя мембрана митохондрий непроницаема для ацил-КоА, образовавшегося в цитоплазме. Переносчиком активированной жирной кислоты является карнитин (у-триметиламино-в-гидроксибутират) (CH3)3N—СН2—СН(ОН)—СН2—СООН. Это широко распространенное соединение, особенно много его в мышечной ткани. В транспорте ацил-КоА принимают участие фермент — карнитин-ацилтрансфераза (Е1) и транспортный белок (карнитин: ацилкарнитин-транслоказа). Ацил-КоА, соединяясь с карнитином, при действии карнитин-ацилтрансферазы образует ацилкарнитин (эфир карнитина и жирной кислоты), который при участии транслоказы проникает внутрь митохондрии. После прохождения ацилкарнитина через мембрану митохондрии происходит обратная реакция — расщепление ацилкарнитина при участии митохондриального HS-KoA и карнитин-ацилтрансферазы (Е1). Карнитин-ацилтрансфераза является основным регуляторным ферментом процесса окисления жирных кислот. Ингибитором этого фермента является исходный интермедиат синтеза жирных кислот — малонил-КоА. Таким образом, если активируется липогенез, увеличивается концентрация малонил-КоА, который ингибирует карнитин-ацил-КоА-трансферазу и выключает р-окисление.
|
|
|
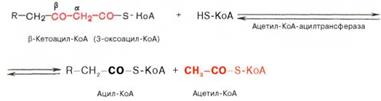

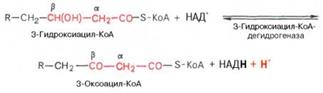
 • Последовательность реакций в-окисления ацил-КоА в матриксе митохондрий. Эта система локализована в матриксе митохондрий в непосредственной близости от дыхательной цепи, интегрированной во внутреннюю мембрану митохондрий. Таким образом, окисление ацил-КоА до ацетил-КоА, в процессе которого происходит восстановление НАД+ и ФАД. 1. Реакция дегидрирования, катализируемая ФАД-зависимой ацил-КоАдегидрогеназой, приводит к образованию а, в-ненасыщенного ацил-SKoA. Фермент обладает стереоспецифичностью, поэтому в результате этой реакции образуется только транс-изомер {транс-еноил-КоА). 2. Во второй реакции происходит гидратация ненасыщенного транс-еноил-КоА при действии фермента еноил-КоА-гидратазы. В результате образуется L-a-в-гидроксиацил-КоА. 3. Реакция дегидрирования, в процессе которой образовавшийся а-в-гидроксиацил-КоА дегидрируется в в-положение. Эту реакцию катализирует НАД-зависимая в -гидроксиацил-КоА-дегидрогеназа. 4. В заключительной реакции тиолитического расщепления в-кетоацил-КоА с помощью еще одной молекулы коэнзима А образуется укороченный на два углеродных атома ацил-КоА и двухуглеродный фрагмент в виде ацетил-КоА. Реакция катализируется ацетил-КоА-ацилтрансферазой (или тиолазой). Образующийся укороченный ацил-КоА вновь вступает в следующий цикл Р-окисления, начиная с первой реакции дегидрирования, и происходит повторное превращение этого ацил-КоА в цикле, состоящем из четырех реакций, и т. д. Такой процесс р-окисления протекает до образования четырехуглеродного соединения — ацетоацетил-КоА. Последняя реакция тиолитического расщепления этого соединения приводит к образованию двух молекул ацетил-КоА и тем самым завершает в целом распад жирной кислоты по механизму в-окисления. Суммарное уравнение окисления активированной пальмитиновой кислоты можно записать следующим образом: СН3(СН2)14СО-КоА + 7ФАД + 7НАД+ + 7Н2O + 7HS-KoA –> 8СН3СО-КоА + 7ФАДН2 + 7НАДН + 7Н+.
• Последовательность реакций в-окисления ацил-КоА в матриксе митохондрий. Эта система локализована в матриксе митохондрий в непосредственной близости от дыхательной цепи, интегрированной во внутреннюю мембрану митохондрий. Таким образом, окисление ацил-КоА до ацетил-КоА, в процессе которого происходит восстановление НАД+ и ФАД. 1. Реакция дегидрирования, катализируемая ФАД-зависимой ацил-КоАдегидрогеназой, приводит к образованию а, в-ненасыщенного ацил-SKoA. Фермент обладает стереоспецифичностью, поэтому в результате этой реакции образуется только транс-изомер {транс-еноил-КоА). 2. Во второй реакции происходит гидратация ненасыщенного транс-еноил-КоА при действии фермента еноил-КоА-гидратазы. В результате образуется L-a-в-гидроксиацил-КоА. 3. Реакция дегидрирования, в процессе которой образовавшийся а-в-гидроксиацил-КоА дегидрируется в в-положение. Эту реакцию катализирует НАД-зависимая в -гидроксиацил-КоА-дегидрогеназа. 4. В заключительной реакции тиолитического расщепления в-кетоацил-КоА с помощью еще одной молекулы коэнзима А образуется укороченный на два углеродных атома ацил-КоА и двухуглеродный фрагмент в виде ацетил-КоА. Реакция катализируется ацетил-КоА-ацилтрансферазой (или тиолазой). Образующийся укороченный ацил-КоА вновь вступает в следующий цикл Р-окисления, начиная с первой реакции дегидрирования, и происходит повторное превращение этого ацил-КоА в цикле, состоящем из четырех реакций, и т. д. Такой процесс р-окисления протекает до образования четырехуглеродного соединения — ацетоацетил-КоА. Последняя реакция тиолитического расщепления этого соединения приводит к образованию двух молекул ацетил-КоА и тем самым завершает в целом распад жирной кислоты по механизму в-окисления. Суммарное уравнение окисления активированной пальмитиновой кислоты можно записать следующим образом: СН3(СН2)14СО-КоА + 7ФАД + 7НАД+ + 7Н2O + 7HS-KoA –> 8СН3СО-КоА + 7ФАДН2 + 7НАДН + 7Н+.
|
|
|
• энергетику окисления жирных кислот. При расчете баланса АТФ в этом процессе следует предположить, что все образовавшиеся молекулы ацетил-КоА включаются в цикл, трикарбоновых кислот и их полное окисление сопровождается синтезом 12 молекул АТФ в расчете на окисление одной молекулы ацетила. НАДН и ФАДН2 окисляются в митохондриальной дыхательной цепи. При окислении НАДН синтезируются 3 АТФ, а ФАДН2 — 2 АТФ и, следовательно, в одном цикле в-окисления синтезируется 5 АТФ. Таким образом, расчет баланса АТФ при полном окислении жирной кислоты с числом углеродных атомов, равным n, можно провести по формуле: (5АТФ*(n/2 -1)+ 12 АТФ*(n/2))-1АТФ, где (n/2 — 1)— число циклов в-окисления; n/2 — число образовавшихся молекул ацетил-КоА; 1 АТФ — затрачивается на активацию жирной кислоты. Следовательно, при окислении 1 молекулы пальмитиновой кислоты (С16) баланс АТФ составляет 130 молекул: [5АТФ-7 + 12АТФ • 8] — 1АТФ = 130 АТФ.
|
|
|
