 |


Теоретическое исследование. Возможности создания биолазера
|
|
|
|
ТЕОРЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
ВОЗМОЖНОСТИ СОЗДАНИЯ БИОЛАЗЕРА
НА ФРЕЛИХОВКИХ МОДАХ [3]
В данной главе обсуждается и аналитически рассматривается возможность создания перевозбужденного состояния основной (выделен-ной) коллективной Фрелиховской моды за счет когерентного резо-нансного взаимодействия электромагнитного (амплитудно-модулиро-ванного) излучения с Фрелиховским осциллятором. В рамках по-нятий лазерной физики речь идет о создании инверсной заселенности между квантовыми уровнями выделенной колебательной моды и, в итоге, о реализации “in vitro-in vivo” суперфлуоресценции и лазерной генерации с использованием в качестве рабочих тел молекул ДНК, РНК, белков, а также таких надмолекулярных структур, как рибосомы, полирибосомы и хромосомы.
Подчеркнем, что в отличие от Фрелиховского подхода, в котором подразумевается квазинеравновесное состояние (колебательная температура выделенной моды превосходит таковую “тепловой бани” Tvib> Teq> 0, т. е. колебания квазиравновесны), в данной работе оценены условия, при которых система рассматриваемых биосубстратов инвертирована (Tvib< 0), что прямо связано с созданием инверсной населенности.
Итак, Фрелиховская мода моделируется двухуровневой квантовой системой (уровень 1 основное состояние, 2 верхнее), возбуждаемой резонансным амплитудно-модулированным электрическим полем
E( t)=Eog(t)сoswt , (1)
где Eo амплитуда напряженности поля, g(t) модуляционный фактор, w=w21 (w21 частота перехода 2®1).
Процесс возбуждения колебаний моды описывается уравнением Больцмана для матрицы плотности:

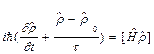 , (2)
, (2)
где оператор гамильтона в дипольном приближении имеет вид: 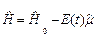
|
|
|
где Ho= 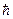 w21
w21 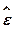 гамильтониан изолированной двухуровневой системы, оператору соответствует матрица с элементами
гамильтониан изолированной двухуровневой системы, оператору соответствует матрица с элементами 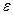 11= 12= 21=0, 22=1, оператор прекции индуцированного дипольного момента осциллятора на направление поля,
11= 12= 21=0, 22=1, оператор прекции индуцированного дипольного момента осциллятора на направление поля, 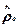
 равновесная матрица плотности,
равновесная матрица плотности, 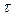 феноменологически введенное время релаксации (для диагональных элементов
феноменологически введенное время релаксации (для диагональных элементов 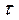 =T1, для недиагональных T2).
=T1, для недиагональных T2).
Уравнению Больцмана (2) эквивалентна следующая система уравнений для элементов матрицы плотности ( 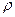 ik; i, k=1, 2):
ik; i, k=1, 2):
i ( 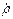 11+( 11 1)/T1)= E(t)(
11+( 11 1)/T1)= E(t)( 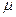 21 12 - 12 21),
21 12 - 12 21),
i ( 12+ 12/T2)= - 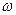 21 12-E(t) 12( 22 - 11), (3)
21 12-E(t) 12( 22 - 11), (3)
i (  21+ 21/T2)= + 21 21+E(t) 21( 22 - 11)
21+ 21/T2)= + 21 21+E(t) 21( 22 - 11)
с учетом уровня нормировки
22+ 11=1 (4)
Нетрудно показать, что система (3) может сводиться к уравнению (при выкладках вторыми гармониками ~exp(2i 21t) пренебрегалось): 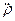 22+
22+ 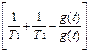 22+
22+ 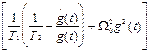
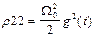
22 (0) = 22 = 0, (5)
где 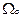 =Eo 21/ частота Раби. Заметим, что амплитудная модуляция поля приводит не только к модуляциям частоы Раби, но и к модуляции “коэффициента трения” осциллятора.
=Eo 21/ частота Раби. Заметим, что амплитудная модуляция поля приводит не только к модуляциям частоы Раби, но и к модуляции “коэффициента трения” осциллятора.
Ниже рассматривается случай T1=T2=T. Можно показать, что уравнение (5) допускает точное решение для произвольной функции g(t):
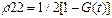 (6)
(6)
G(t)= 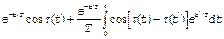

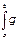 (t’)dt’ (7)
(t’)dt’ (7)
Рассмотрим случай периодической модуляции амплитуды напряженности поля
g(t)=cos 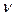 t. (8)
t. (8)
Если период модуляции T 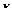 =2
=2 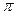 / короче времени релаксации (T
/ короче времени релаксации (T 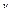 < < T), то для времени T < < t< < T усреднение (6) за период T дает:
< < T), то для времени T < < t< < T усреднение (6) за период T дает:
< 22> =1/2 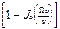 (9)
(9)
и, соответственно, (4):
< 11> =1/2 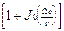 ,
,
где  функция Бесселя нулевого порядка, так что для разности населенностей уровней 2 и 1 имеем
функция Бесселя нулевого порядка, так что для разности населенностей уровней 2 и 1 имеем
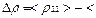
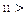 =
= 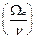 . (10)
. (10)
Из (10) четко следует, что в диапазонах параметра 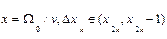 , где k=1, 2,.. и
, где k=1, 2,.. и 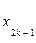 корни функции Бесселя, вероятность заселения уровня 2 превосходит таковую для уровня 1. Другими словами, мы имеем перевозбужденное инвертированное состояние осциллятора, что является необходимым условием для создания условий лазерной генерации (
корни функции Бесселя, вероятность заселения уровня 2 превосходит таковую для уровня 1. Другими словами, мы имеем перевозбужденное инвертированное состояние осциллятора, что является необходимым условием для создания условий лазерной генерации ( 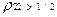 ). Ситуация здесь аналогична процессу раскачивания маятника с пульсирующей точкой подвеса (маятник Капицы, классическое рассмотрение[29]).
). Ситуация здесь аналогична процессу раскачивания маятника с пульсирующей точкой подвеса (маятник Капицы, классическое рассмотрение[29]).
|
|
|
Для больших времен, t> > T, функция G(t), входящая в соотношение (6), имеет вид:
G(t)=P(t)cos  + Q(t)sin
+ Q(t)sin 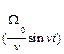 ,
,
P(t)= 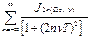
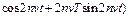
Q(t)=2 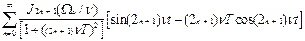 , (11)
, (11)
где J функция Бесселя соответствующего порядка.
Из (11) следует важный вывод: когерентный механизм взаимодействия Фрелиховских мод с резонансным амплитудно-модулированным полем обусловливает незатухающие колебания диагональных элементов матрицы плотности для времен t, превосходящих времена релаксации системы, причем частоты пульсаций кратны частоте амплитудной модуляции .
Усредняя (11) за период T , получаем
< G(t)> = 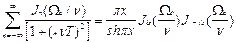 , (12)
, (12)
где x= 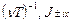 функции Бесселя мнимого порядка (i мнимая единица). В частном случае, когда период модуляции T короче времени релаксации T, x < < 1,
функции Бесселя мнимого порядка (i мнимая единица). В частном случае, когда период модуляции T короче времени релаксации T, x < < 1,
< 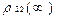 > =1/2
> =1/2 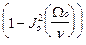 , <
, < 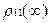 > =1/2
> =1/2 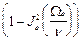 , (13)
, (13)
так что
< 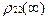 > <
> < 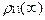 > = -
> = - 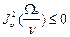 . (14)
. (14)
В данном случае эффект инверсии не реализуется.
Рассмотрим случай, когда закон модуляции задается соотношением
g(t)=1+ 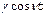 . (15)
. (15)
По аналогии с предыдущим для функции G(t), входящей в соотношение (6), можно получить (T 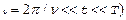 .
.
G(t)= 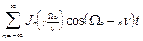 . (16)
. (16)
Из (16) видно, что спектр пульсаций диагональных матричных элементов 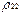 и
и 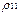 включает, кроме частоты Раби, “стоксовые” и “антистоксовые” комбинационные частоты
включает, кроме частоты Раби, “стоксовые” и “антистоксовые” комбинационные частоты 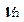
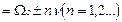 . Допустим для определенных n выполнено условие
. Допустим для определенных n выполнено условие 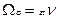 , т. е.
, т. е.
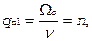 (17)
(17)
тогда, как следует из (16), постоянная составляющая для вероятностей 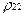 и
и 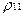 сдвигается. Динамическому состоянию равновесия при этом соответствуют величины:
сдвигается. Динамическому состоянию равновесия при этом соответствуют величины:
< > =1/2 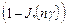 , <
, < 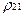 > =1/2
> =1/2 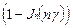 , (18)
, (18)
так что 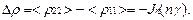
Эффект инверсии ( 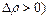 реализуется при условии
реализуется при условии
 . (19)
. (19)
Если параметр глубины модуляции 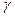 лежит в диапазонах, где значения функции Бесселя
лежит в диапазонах, где значения функции Бесселя  отрицательны, то реализуется режим перевозбуждения системы (информационных биомакромолекул и надмолекулярных структур).
отрицательны, то реализуется режим перевозбуждения системы (информационных биомакромолекул и надмолекулярных структур).
Таким образом, высказана идея принципиальной возможности создания биолазеров на Фрелиховских модах in vitro, а также инициации таких процессов в живой клетке в дополнение (или коррекции) к известным естественным лазероподобным процессам в биосистемах. Показано, что в определенных условиях в случае когерентного (резо-нансного) взаимодействия амплитудно-модулированного внешнего электромагнитного излучения с Фрелиховской модой система информационных биоструктур может существовать в перевозбужденном состоянии, что является необходимой предпосылкой для создания знаконесущих биолазеров.
|
|
|
Необходимо отметить, что описанный выше механизм формирования биолазеров на основе молекул ДНК позволяет подойти к попытке реализации еще одной фундаментальной гипотезы Фрелиха о возможности перекачки энергии kТ внутриклеточной жидкости в энергию электрических колебаний в молекуле ДНК[30]. В соответствии с этой гипотезой стохастические тепловые колебания kТ раствора могут резонансно взаимодействовать (в определенном интервале частот) с колебательными модами молекулы ДНК, и благодаря тому, что как молекула ДНК, так и молекулы белков представляют собой распределенные нелинейные колебательные структуры, часть энергии может группироваться в низкочастотных модах этих молекул. Иными словами, молекула ДНК в растворе может частично преобразовывать энергию колебаний kТ в энергию собственных мод. Заметим, что даже в рамках предложенного квазили-нейного подхода проблема перекачки тепловой энергии раствора может быть сведена к механизму затухания квантового осциллятора, который был предложен А. Пиппардом[31]. C учетом этого в уравнение Шредингера вводится комплексный потенциал, интерпретирующий передачу энергии осциллятора большому числу мод расширяющегося сферического резонатора. Если размеры этого резонатора конечны, как в случае с живой клеткой, то возникнет резонансный обмен энергии между модами kТ раствора и электрическими модами молекулы ДНК. Эти рассуждения также говорят в пользу того, что и в водно-жидкокристаллическом электролите клеточно-тканевого пространства биосистемы генетические молекулы могут функционировать как биолазеры.
Надо указать на существенное обстоятельство относительно принципиальной возможности реализации возбуждения Фрелиховских мод “in vitro” по биохимическому пути, а именно за счет энергии гидролиза АТФ и других нуклеозид-трифосфатов, а также за счет других макроэргических соединений живой клетки. В данном случае мы будем искусственно повторять то, что эволюционно и (или) иным путем дано биосистемам как основная информационная и, может быть, энергетическая фигура. Эта часть наших исследований ставит определенные нравственные и этические проблемы применения биолазеров.
|
|
|
АНТЕННАЯ МОДЕЛЬ
ФИЗИКО-МАТЕМАТИЧЕСКИЙ ФОРМАЛИЗМ [16]
Как уже неоднократно отмечалось, функционирование ряда биологических макромолекул (в частности, ферментов) и других биологических соединений во многом определяется процессами, происходящими в активных центрах, окруженных биополимерными цепочками, имеющими знаковую топологию. Исходя из такого представления о структуре информационных биомакромолекул, естественно предположить, что их взаимодействие с физическими полями внешних по отношению к биосистеме и внутренних (организменных) излучений приводит к возбуждению дипольно-активных колебаний мономеров, формирующих указанную цепочку, а те, в свою очередь, индуцируют колебания в активном центре. Иными словами, такая система будет работать как своеобразная антенна. Эти возбужденные колебания способны привести к переходу биомакромолекулы в другое конформационное (топологическое, знаковое) состояние.
Подобная концепция в принципиальном плане адекватна целому ряду функционально высокозначимых биомакромолекул, например, хлорофилла, гемоглобина, миоглобина и т. д. Эти макромолекулы объединяются двумя структурными качествами: 1) в их геометрическом центре расположен ион (в случае хлорофилла - ион магния, в случае гемоглобина - ион железа); 2) около иона симметрично расположены 4 пиррольных кольца (псевдоплоская структура).
Другими типами биополимеров, соответствующих антенной модели, могут быть cравнительно простые циклы типа валиномицина (переносчик ионов калия) и сложные надмолекулярные структуры хромосом, ДНК которых содержит высокоорганизованные ассоциаты таких металлов, как магний, кальций, никель, кобальт, медь, железо, цинк и др. При этом роль их неясна и сводится исследователями, в основном, к нейтрализации ОН-групп остатков фосфорной кислоты полинуклеотида. Представляется, что функции металлов в ДНК и РНК существенно более широкие и реализуются по линии знакового и (или) энергетического взаимодействия с эндогенными и экзогенными по отношению к биосистеме физическими полями. То же относится и к белкам, не содержащим порфириновый центр, но специфическим образом связывающим металлы. Например, таковыми можно считать сайт-специфические белки с доменами типа “цинковых пальцев”, участвующими в регуляции генов, подчас очень далеко отстоящих от этих управляющих белков. Атомы металлов ДНК и белков могут резонансно взаимодействовать по электромагнитным каналам в рамках понятий антенной модели. Еще раз обозначим понятие антенной модели.
|
|
|
Внешняя энергия (в частности, связанная с резонансным взаимодействием крайне высокочастотных электромагнитных излучений с белками) поступает на периферию, т. е. на ансамбль субъединиц (не обязательно идентичных по структуре). В результате активной “беседы”, предопределенной биохимическими связями, между периферийными акцепторами (получившими закодированную энергию) и центром-ассоциатом (в данном случае ионом металла гемсодержащих белков), последний получает энергию (информацию), что и вызывает биологическое действие. Степень реакционной способности биомакромолекул существенно зависит от уровня возбуждения центральных субъединиц. Рассмотрим в деталях потенциальные механизмы волновых взаимодействий физических полей и активных центров информационных биомакромолекул в рамках предлагаемой нами антенной модели.
В качестве простейшей модели для иллюстрации антенного эффекта рассмотрим двумерную замкнутую (циклическую) цепочку мономеров. В центре цикла расположен активный центр, связанный с мономерами цепочки диполь-дипольным взаимодействием.
Обозначим координатные смещения мономеров через 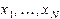 , а смещение активного центра через
, а смещение активного центра через 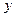 . Для потенциальной функции имеем:
. Для потенциальной функции имеем:
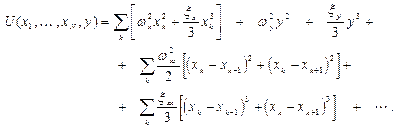 (1)
(1)
Первые два члена в (1) соответствуют колебаниям мономеров (второй член учитывает ангармонизм); последние два члена отвечают за связи между мономерами, Остальные члены отвечают за связи между мономерами и активным центром.
Уравнения движения запишем в виде:
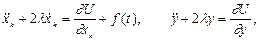 (2)
(2)
где 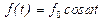 внешняя монохроматическая сила, действующая только на мономеры,
внешняя монохроматическая сила, действующая только на мономеры,  коэффициент затухания, введенный феноменологически (простоты ради принят одинаковым и для мономеров, и для активного центра).
коэффициент затухания, введенный феноменологически (простоты ради принят одинаковым и для мономеров, и для активного центра).
С учетом (1), система уравнений (2) приобретает вид:
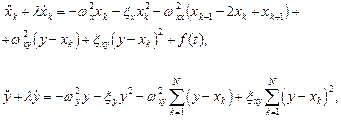 (3)
(3)
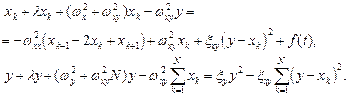 (4)
(4)
Введем общую координату для ансамбля мономеров
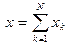 . (5)
. (5)
тогда система уравнений (4) в линейном приближении приобретает вид:
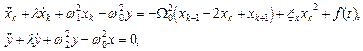 (6)
(6)
где:
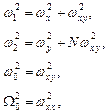
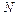 число мономеров.
число мономеров.
С учетом (5) имеем 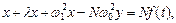 (7. 1)
(7. 1)
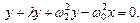 (7. 2)
(7. 2)
Из (7. 2) следует 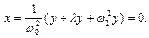 (8)
(8)
Подстановка (8) в (7. 1) дает
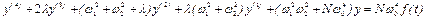 .
.
(9)
Соответствующее характеристическое уравнение имеет вид (после подстановки 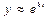 в однородное уравнение):
в однородное уравнение):
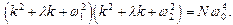 (10)
(10)
Обозначив 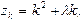 имеем
имеем
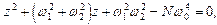
так что
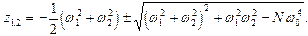 (11)
(11)
В дальнейшем предполагается выполнение неравенств:
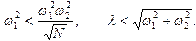 (12)
(12)
Первое условие соответствует случаю слабой связи между мономерами и активным центром, второе - малому затуханию мономерных осцилляторов.
Для собственных значений имеем
 , (13)
, (13)
где введены коллективные частоты:
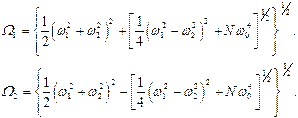 (14)
(14)
Нас интересуют вынужденные колебания (внешняя сила 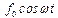 ):
):
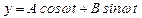 . (15)
. (15)
Подстановка (15) в (9) и приравнивание соответствующих коэффициентов при 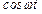 и
и 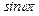 дают систему алгебраических уравнений:
дают систему алгебраических уравнений:
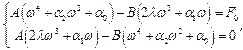
где:
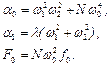
В результате получаем 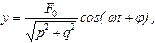
где 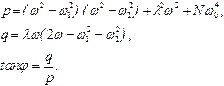
После несложных, но громоздких преобразований для вынужденных колебаний активного центра получаем:
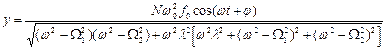 . (16)
. (16)
Из (16) видно, что наибольшая амплитуда вынужденных колебаний активного центра достигается в условиях коллективного резонанса: либо 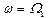 , либо
, либо 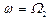 .
.
В любом из этих случаев для амплитуды вынужденных колебаний имеем:
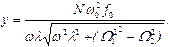 (17)
(17)
Из (17) следует, что наибольший эффект резонансной раскачки активного центра достигается при большем числе периферийных субъединиц “антенны”, при более высоком значении коэффициента связи активного центра с мономерами, при наименьшем коэффициенте затухания и при наименьшем дисбалансе коллективных мод.
Нетрудно определить и “хореографию” (динамику вынужденных колебаний) отдельных мономерных единиц. В соответствии с (6) уравнение для k -го мономера запишем в виде:
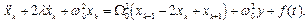 (18)
(18)
Вводя коллективные координаты
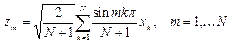
и применяя метод линейной алгебры, получаем для вынужденных колебаний мономеров:
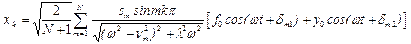 ,
,
(19)
где:

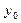 ‑ определяется из (16).
‑ определяется из (16).
Таким образом, в рамках антенной модели наибольший эффект воздействия внешнего монохроматического поля 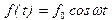 ре-ализуется в условиях коллективного резонанса:
ре-ализуется в условиях коллективного резонанса:
 .
.
Повторяя рассуждения раздела 2, можно сделать также следующие выводы:
1) При реализации амплитудной модуляции внешнего сигнала имеют место дополнительные возможности резонансного воздействия на биомакромолекулы на частотах:
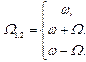
2) Учет нелинейности при квадратичной связи для монохроматического сигнала привносит дополнительный резонанс на второй гармонике 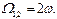
3) Учет нелинейности при амплитудной модуляции определяет еще ряд резонансных возможностей:
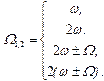
|
|
|
