 |


Рис. 46. Сплайсинг пре-мРНК (цит. по Северину, 2002)
|
|
|
|
Рис. 46. Сплайсинг пре-мРНК (цит. по Северину, 2002)
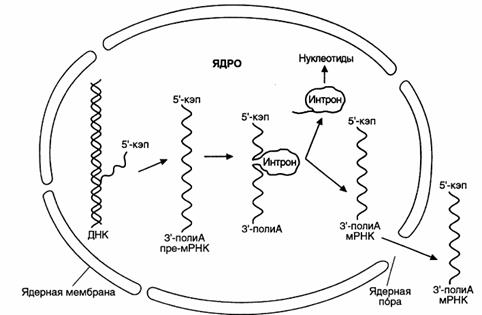
Рис. 47. Образование и выход из ядра зрелой мРНК (цит. по Северину, 2002)
 |
Рис. 48. Альтернативный сплайсинг (цит. по Северину, 2002)
После завершения процессинга зрелые мРНК перемещаются в цитоплазму при помощи специальных белков-информоферов.
Посттранскрипционные модификации тРНК
тРНК синтезируются в виде 4, 5S РНК-предшественников (~100 нуклеотидов). В ядре при формировании пространственной конформации зрелых молекул (рис. 49) пре-тРНК укорачиваются с 5'- и 3'-концов и вырезается один интрон из центральной части полинуклеотидной цепи с помощью специфических РНКаз (реакция 1).
Модифицируются азотистые основания и к 3’-концу тРНК с помощью нуклеотидилтрансферазы последовательно присоединяется триплет ССА (реакция 2).
Зрелые молекулы тРНК выходят из ядра в цитоплазму (реакция 3).

Рис. 49. Созревание пре-тРНК (цит. по Северину, 2002)
Посттранскрипционные модификации рРНК
Пре-рРНК освобождаются из комплекса с ДНК в виде крупного транскрипта с константой седиментации 45S, который является предшественником 18S, 28S и 5. 8S рРНК.
1-2% нуклеотидов этой молекулы метилируется по 2'-гидроксильной группе рибозы, т. о.
«отмечаются» участки, которые должны войти в состав зрелых рРНК. Самая короткая 5S рРНК кодируется отдельным геном, транскрибируется РНК-полимеразой III (ответственной за синтез тРНК) и затем поступает в 60 S рибонуклеопротеиновую частицу.
Субъединицы рибосомы и все зрелые мРНК и тРНК поступают в цитоплазму.
Исследования, проводимые на кафедре биохимии КГУ, связанные с обменом нуклеиновых кислот
В 1980х гг. была открыта новая функция антител – способность катализировать различные биохимические реакции, то есть выступать в роли биологических катализаторов. По аналогии с энзимами (англ. enzyme) такие антитела были названы абзимы (от англ. a nti b ody en zyme ), или каталитические антитела. Согласно сложившимся представлениям, наличие в крови абзимов является чётким признаком протекания в организме аутоиммунных процессов. Но механизмы реализации патологических свойств абзимов и их клиническое значение пока остаются невыясненными.
|
|
|
Под научным руководством проф. Винтера В. Г. была инициирована и разработана одна из основных плановых тем кафедры биохимии КГУ – изучение ферментативной активности и биологической роли антител к нуклеиновым кислотам, обладающих ДНК-гидролизующей активностью, при различных заболеваниях человека. Не исключено, что природные абзимы могут быть антителами с уникальным гидролитическим центром. Учитывая тот факт, что
антитела к ДНК проникают в клетку и ядро, ДНК-гидролизующие антитела могут участвовать в репликации, репарации и рекомбинации ДНК, вмешиваясь в ход событий этих процессов, что приводит к изменению пролиферации и апоптоза клеток. Активность абзимов к ДНК может модулироваться состоянием клетки.
Следовательно, изучение взаимодействия антител с нуклеиновыми кислотами и ферментативной активности абзимов к ДНК позволит выяснить не только фундаментальные основы функционирования иммунной системы, но и внесет вклад в понимание биохимических основ жизнедеятельности клетки.
ТЕСТЫ ПО ТЕМЕ “ОБМЕН НУКЛЕИНОВЫХ КИСЛОТ”
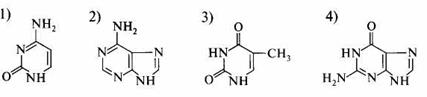
1. К пиримидиновым основаниям относится:
 |
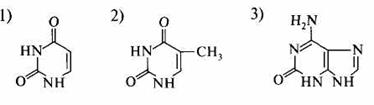
2. К пуриновым основаниям относится:
 |
3. Входит в состав:
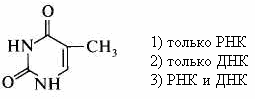 |
4. Входит в состав:
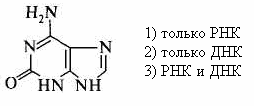 |
5. Является:
6.  Является:
Является:

7. В продуктах полного гидролиза нуклеиновых кислот отсутствуют:
|
|
|
1) азотистые основания
2) пентозы
3) гексозы
4) фосфорные кислоты
8. В нуклеотидах азотистое основание и пентоза соединены связью:
1) фосфоангидридными
2) 2', 3'-фосфодиэфирными
3) 3', 5'-фосфодиэфирными
4) 2', 5'-фосфодиэфирными
5) N-гликозидными
9. Модель вторичной структуры ДНК предложена:
1) Р. Митчелом и В. П. Скулачевым
2) Дж. Уотсоном и Ф. Криком
3) Ф. Жакобом и Ж. Моно
|
|
|
