 |


Методические рекомендации по самоподготовке
|
|
|
|
Тема «ГУМОРАЛЬНАЯ РЕГУЛЯЦИЯ ФУНКЦИЙ.
ФИЗИОЛОГИЯ ЖЕЛЁЗ ВНУТРЕННЕЙ СЕКРЕЦИИ»
(1-2 занятие)
- Продолжительность изучения темы: 8 часов;
из них на занятие 6 часа; самостоятельная работа 2 часа
- Место проведения: учебная комната
Цель: а) знать структурно-функциональную организацию эндокринной системы, образование, выделение, перенос и распад гормонов, их основные механизмы действия, саморегуляцию эндокринной системы, функциональные связи желез внутренней секреции и нервной системы, нейросекреты гипоталамуса; основные виды влияний гормонов на органы и системы организма; методы исследования эндокринной функции;
б) уметь оценивать роль эндокринных механизмов в регуляции гомеостаза, адаптации организма к условиям внешней среды, в процессах высшей нервной деятельности и поведения.
Мотивация темы: гормональная регуляция является обязательным элементом поддержания гомеостаза, участвует в реализации высших психических функций: мотивации, эмоции, сна, памяти. Адаптивные реакции организма, в том числе стресс, протекают при обязательном участии гормонов. Нарушения функции эндокринных желез являются одной из наиболее распространенных форм патологии, с которой сталкивается врач любой специальности. Знания в области эндокринологии необходимы как для своевременной и точной диагностики
Методические рекомендации по самоподготовке
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логическую структуру учебного материала.
Эндокринная система - совокупность взаимосвязанных и различным образом организованных эндокринных клеток, осуществляющая централизованное управление функций организма с помощью сигнальных молекул - гормонов. Гормоны - внеклеточные сигнальные регуляторы, образующиеся специализированными эндокринными клетками и обладающие дистантным действием..
|
|
|
Виды объединения эндокринных клеток.
- Формируют эндокринные железы.
- Образуют часть железистого (например, островки поджелудочной железы, гонады) или нежелезистого (например, почки, сердце) органа.
- Эндокринные клетки диффузно расположены в органах (гастроинтестинальная система).
- Образование эндокринной системы как результат взаимодействия эндокринных структур.
Виды действия гормонов.
Эндокринное (дистантное) действие - гормоны эндокринной железы действуют на клетки-мишени других органов и тканей.
Паракринное действие - гормоны эндокринной клетки действуют на соседние клетки в пределах одного органа (например, гастроинтестинальные гормоны, простагландины).
Аутокринное действие - гормоны эндокринной клетки действуют на эту же клетку (например, простагландины).
Химическая характеристика гормонов.
Основные классы гормонов.
1.Белково-пептидные гормоны (например, гормоны гипофиза).
2. Производные аминокислот (из тирозина образуются адреналин, норадреналин, дофамин, тироидные гормоны, из триптофана - мелатонин, серотонин).
3. Стероидные гормоны (кортикостероиды, половые гормоны, 1,25-(ОН)2-витамин D3).
Основные фрагменты молекулы гормонов.
Адресный фрагмент обеспечивает связывание гормона с циторецептором например, α -субъединица лютеинизирующего гормона. (Снижение этого сродства приводит к гормонорезистентности.) Актон обеспечивает специфичность действия гормона (например,β - субъединица лютеинизирующего гормона). Вспомогательные фрагменты служат для изменения его действия, определяют его иммунологические свойства.
Секреция гормонов.
|
|
|
Экзоцитоз секреторных гранул (пептидные гормоны, катехоламины) стимулируется Са2+, поступившим в цитозоль через Са2+-каналы при деполяризации мембраны клетки или (и) из гладкой ЭПС при действии инозитол-трифосфата.
Освобождение гормона из белковосвязанной формы (тироксин из иодированного тироглобулина).
Свободная диффузия гормонов, растворимых в липидах мембраны (например, кортикостероиды, половые гормоны).
Циркуляторный транспорт гормонов (основные формы их транспорта в крови).
Свободная, несвязанная форма составляет от сотых долей процента (например, Т4 и Т3) до 100 % (например, инсулин). Только эта форма гормона действует на клетки-мишени.
Специфическое связывание с глобулинами (70 - 80 %) обеспечивает создание запаса и связывание избытка гормона в крови, защиту его от инактивации и от потери через почки. (Виды глобулинов - транскортин, тироксин-связывающий глобулин, прогестин, тестостерон-эстрогенсвязывающий глобулин.)
♦ Неспецифическое связывание с альбуминами крови (5 - 10 %). Благодаря низкой прочности связи они являются основным источником свободных гормонов в крови. (Небольшая часть гормона связана с форменными элементами крови.)
♦ Периферическая конверсия гормонов - превращение гормонов в органах и тканях, отдаленных от эндокринной железы, в результате которого изменяется активность гормона (например, превращение тироксина в трииодтиронин в печени, тестостерона в дигидротестостерон - в простате).
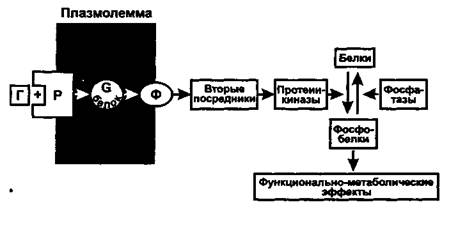
Рис. 1. Общая схема действия гормонов через вторые посредники. Г - гормон, Р - рецептор, Ф - фермент
Физиологические эффекты гормонов.
Клетки - (ткани-, органы-) мишени, избирательно реагирующие на гормоны, имеют циторецепторы - белковые приемники и преобразователи гормонального сигнала. (Есть данные, что на один рецептор может действовать более, чем один гормон.) Циторецепторы расположены на клеточной мембране или в ядре (500-30000 рецепторов в клетке). Максимальный эффект гормона наблюдается при его концентрации, обеспечивающей занятость всего 5 - 10 % общего числа рецепторов (феномен «резервных» рецепторов). Изменение числа рецепторов является способом регуляции гормонального ответа (активация и ингибирование синтеза рецепторов, интернализация рецепторов путем эндоцитоза).Гормоны, плохо проникающие в клетку (белково-пептидные, катехоламины), соединяются с рецепторами на клеточной мембране и вызывают внутриклеточные эффекты через вторые посредники. Большинство гормонов после присоединения к рецепторам активируют регулируемый гуанилнуклеотидами С-белок мембраны. Имеется несколько видов G-белков (Gs, Gi, Gq, G0), которые влияют на ферменты (например, аденилатциклазу, фосфолипазу С), образующие различные вторые посредники (например, цАМФ, инозитолтрифосфат, диацилглицерол). При этом G-белки могут как активировать (Gs), так и ингибировать (GO образование вторых посредников. Вторые посредники, в свою очередь, активируют соответствующие им протеинкиназы, фосфорилирующие различные клеточные белки по остаткам серина и треонина, что изменяет их функциональную активность (чаще увеличивает). Наряду с этим активированные G-белки оказывают выраженное влияние на проницаемость кальциевых и калиевых каналов мембраны.В действии гормонов, плохо проникающих в клетки, можно выделить 4 системы: аденилатциклазную, фосфоинозитидную, гуанилатциклазную и тирозинкиназную.
|
|
|

Рис. 2 Схема действия гормонов через аденилатциклазную систему. Г - гормон, Р - рецептор, Ф - фермент, ФДЭ - фосфодиэстераза.
• Аденилатциклазная система. Эту систему используют гормоны, осуществляющие свой эффект через аденилатциклазу и синтез второго посредника циклического АМФ (3,5-цАМФ). Действие гормона может как активировать, так и тормозить его образование.
Комплекс «гормон + рецептор» активирует Gs-белок, который активирует аденилатциклазу и образование цАМФ (например, так действуют адреналин через рУр2-рецепторы, АДГ через У2-рецепторы, глюкагон, АКТГ, ТТГ, ФСГ, ЛГ, МСГ, кортиколиберин, соматолиберин, кальцитонин, хориогона-дотропин человека, паратгормон и др.).
Комплекс «гормон + рецептор» активирует Gi-белок, который тормозит аденилатциклазу и образование цАМФ (например, так действуют адреналин через а2-рецепторы, ангиотензин II через АТ-рецепторы, аденозин через Ai-рецепторы, соматостатин, опиоиды через, 5- и к-рецепторы, ацетилхолин через Мг-холинорецепторы гладкомышечных клеток).
|
|
|
цАМФ влияет на функции клетки через изменение активности протеинфосфокиназы-А, фосфорилирующей различные клеточные белки: мембранные (изменяется ее проницаемость), ферменты (изменяется метаболизм клетки), ядерные (изменение транскрипции), рибосом (изменение трансляции), микротрубочек (двигательные эффекты, изменение формы клетки) и др.
• Фосфоинозитидная система. Эту систему используют гормоны, которые, действуя через Оч-белок, активируют в мембране фосфолипазу С. Она образует из фосфолипида мембраны (фосфатидилинози-толдифосфата) два посредника - инозитол-1,4,5-трифосфат (ИФз) и 1,2- диацилглицерол (ДАГ).

Рис. 3. Схема действия гормонов через фосфоинозитидную систему. Г - гормон, Р - рецептор, ФЛ-С - фосфолипаза С, ФИФ2 - фосфатиди-линозитолдифосфат, ИФ3 - инозитолтрифосфат, ДАГ - диацилглицерол
ИФз, переходя в цитоплазму, действует на рецепторы гладкой ЭПС, что приводит к открыванию Са2+-каналов и выходу Са2+ в цитозоль. Са2+ при этом выступает в качестве третьего посредника, который связывается с белком кальмодулином и активирует Са2+-кальмоду-линзависимую протеинкиназу, фосфорилирующую эффекторные белки, влияющие на функции клеток, их пролиферацию и дифференцировку (регулирует активность аденилатциклазы и синтез цАМФ, фосфолипазы-Аг и синтез простагландинов, тромбоксана, простациклина. гуанилатциклазы и синтез цГМФ, фосфодиэстеразы и разрушение цАМФ и цГМФ; участвует в регуляции секреции инсулина, гормонов щитовидной железы, гипофиза и надпочечников, кишечной секреции и секреции нейромедиаторов). (Роль Са2+ как посредника действия гормонов обусловлена его сильным влиянием при низких концентрациях на метаболизм и функции клетки. Гормоны могут увеличивать концентрацию Са2+ в цитозоле как в результате открывания Са2+-каналов клеточной мембраны, так выхода Са2+ из гладкой ЭПС. В клетке есть специальные белки, связывающие Са2+ при низкой концентрации -кальмодулин, гуанилатциклаза, тропонин С и др. Присоединение Са2+ к этим белкам изменяет их конфигурацию и свойства. Это приводит к изменению многочисленных функций клеток - метаболической, секреторной, сократительной, биоэлектрической и др.
ДАГ, оставаясь на внутренней поверхности мембраны, активирует протеинкиназу С, фосфорилирующую эффекторные белки (например, активирует киназу легкой цепи миозина), а также синтез простагландинов из арахидоновой кислоты, что изменяет функции клетки.
Через ИФз и ДАГ действуют адреналин (oci -рецепторы), вазопрессин (Vi), ацетилхолин, ангиотензин П, гистамин, холецистокинин, окситоцин, ВИП и др.
|
|
|
• Гуанилатциклазная система. Некоторые гормоны (например, ацетилхолин, цитокины) действуют через гуанилатциклазу, образующую второй и посредник - циклический ГМФ (цГМФ). Гуанилатциклаза существует в двух формах - мембраносвязанной и растворимой в цитозоле. Мембраносвязанная гуанилатциклаза является частью рецептора плазмолеммы, ее активация гормоном (например, На+- уретическим) увеличивает образования цГМФ, который стимулирует протеинкиназу G, фосфорилирующую белки. Растворимая в цитозоле гуанилатциклаза содержит в своем составе гем, с которым связывается сигнальная молекула - NО (оксид азота образуется из аргинина в ряде клеток - эндотелии, нейронах и др. под действием ацетилхолина, механических раздражителей и др.). Активированная оксидом азота гуанилатциклаза через протеинкиназу G фосфорилирует белки (например, так действует ацетилхолин, вызывая расширение сосудов).
Тирозинкиназная система характеризуется особыми рецепторами, которые путем фосфорилирования и дефосфорилирования сигнальных белков по тирозиновым остаткам, регулируют процессы роста, дифференцировки, выживания, метаболизма и смерти клеток. Эту систему используют некоторые гормоны - инсулин, инсулиноподобные факторы роста (или соматомедины), факторы роста эпидермиса, тромбоцитов и др. Многие рецепторы к таким гормонам содержит в своем составе тирозинкиназу, которая при действии гормона аутофосфорилирует внутреннюю часть рецептора по остаткам тирозина. Активированный таким образом рецептор передает через сложный и недостаточно изученный комплекс посредников фосфорилирующее и дефосфорилирующее влияние по остаткам тирозина на многие сигнальные молекулы цитоплазмы и ядра. Фосфорилирование и дефосфорилирование белков может приводить как к их активации, так и инактивации.
В действительности гормоны оказывают преимущественно разветвленные эффекты. Это связано с тем, что комплекс Г+Р действует с разной степенью на несколько G-белков. В свою очередь активированные G-белки действуют на несколько эффекторов (ферментов, каналов).
Клеточные эффекты липорастворимых гормонов, легко проникающих через клеточные мембраны (стероидные и тироидные гормоны, 1,25-(ОН)г-витамин D3). Гормон соединяется с ядерным рецептором. Рецепторы представляют собой белки (факторы транскрипции), прочно связанные с ДНК хроматина и дополнительными ядерными белками. Каждый рецептор содержит несколько специфических участков (доменов): ДНК-связывающий домен, гормонсвязывающий домен (гидрофобный «карман»), домен, активирующий транскрипцию и др. Активируются процессы транскрипции (образование мРНК с помощью фермента РНК-полимеразы II) и трансляции (взаимодействие рРНК, тРНК и мРНК, индуцирующее синтез белков и дифференцировку клеток). Например, глюкокортикоиды стимулируют гены, обеспечивающие синтез.
|
|
|
