 |


Липогенез.
|
|
|
|
АТФ необходим для синтеза активных форм субстратов, используемых в процессе синтеза жира.
Для синтеза нейтрального жира необходим глицерин в активной форме - глицерол-3-фосфат (фосфоглицерин). Глицерол-3-фосфат может быть получен двумя способами:
1. Путем активации глицерина с помощью глицеринкиназы.
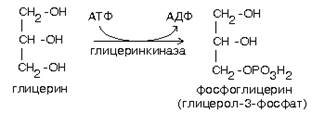
2. Путем восстановления фосфодиоксиацетона, полученного при распаде глюкозы.

Кроме глицерина, для синтеза нейтрального жира необходимы жирные кислоты в активной форме. Активная форма любой жирной кислоты – Ацил-КоА. Образуется при участии фермента ацил-КоА-синтазы.

Здесь наблюдается глубокий распад АТФ до АМФ. АМФ не может вступить в окислительное фосфорилирование. Поэтому существует реакция: АТФ + АМФ Þ 2АДФ. Поэтому затраты на активацию молекулы жирной кислоты эквивалентны затрате двух АТФ. Следующим этапом на пути синтеза жира является реакция образования фосфатидной кислоты:
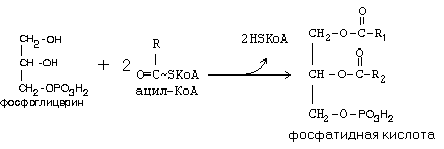
Реакция катализируется ключевым ферментом липогенеза – глицерол-3-фосфатацилтрансферазой. Для этого фермента нет аллостерических эффекторов, но обнаружен адипсин (ацилстимулирующий белок), который облегчает взаимодействие Ацил-КоА с ферментом. Адипсин является продуктом протеолиза одного из компонентов системы комплемента. Относится к гормонам местного действия, так как вырабатывается в жировой ткани и действует там же.
Две последующие реакции являются завершающими в синтезе триацилглицерина.

Реакции синтеза не зависят от того, каково происхождение веществ – участников реакций.
Жир может синтезироваться как из продуктов распада жира, так и из углеводов. Синтез эндогенного жира из углеводов протекает в печени и в жировой ткани. Ацил-КоА синтезируется из Ацетил-КоА. ГБФ-путь распада углеводов обеспечивает синтез энергией. Образование Ацетил-КоА происходит в матриксе митохондрий. Синтез жирных кислот идет в цитоплазме на мембранах эндоплазматического ретикулума путем постепенного удлинения ацетил-КоА на 2 углеродных атома за каждый цикл. Удлинение высших жирных кислот, содержащих более 16 углеродных атомов, идёт путём реакций, обратных b-окислению (О b-окислении смотрите ниже, раздел «Катаболизм жирных кислот»). Однако реакции синтеза жирных кислот до 16 углеродных атомов принципиально отличаются от реакций, обратных b-окислению. Они протекают обходным обратным путём.
|
|
|
Отличия реакций синтеза высших жирных кислот от обратных бета-окислению:
1. b-окисление протекает в митохондриях, а синтез жирных кислот протекает в цитоплазме на мембранах эндоплазматического ретикулума. Но образуется Ацетил-КоА в митохондриях, а через мембраны сам проходить не может. Поэтому существуют механизмы транспорта Ацетил-КоА из митохондрий в цитоплазму.

Ключевыми ферментами ЦТК являются цитратсинтаза и изоцитратдегидрогеназа. Основные аллостерические регуляторы этих ферментов - это АТФ и АДФ. Если в клетке много АТФ, то АТФ выступает как ингибитор этих ключевых ферментов. Однако изоцитратдегидрогеназа угнетается АТФ сильнее, чем цитратсинтетаза. Это приводит к накоплению цитрата и изоцитрата в матриксе митохондрии. При накоплении цитрат выходит из митохондрии в цитоплазму. В цитоплазме есть фермент цитратлиаза. Этот фермент расщепляет цитрат на ЩУК и Ацетил-КоА.
Условием для выхода Ацетил-КоА из митохондрии в цитоплазму является хорошее обеспечение клетки АТФ. Если АТФ в клетке мало, то Ацетил-КоА расщепляется до СО2 и Н2О.
2. В ходе b-окисления промежуточные продукты связаны с HS-КоА, а при синтезе жирных кислот промежуточные продукты связаны с особым ацил-переносящим белком (АПБ). Это сложный белок. Его небелковая часть похожа по строению на КоА. Небелковая часть АПБ состоит из тиоэтиламина, пантотеновой кислоты (витамин В3) и фосфата.
|
|
|
3. При b-окислении в качестве окислителя используются НАД и ФАД. При синтезе ЖК нужен восстановитель - используется НАДФ*Н2.
В клетке существует 2 основных источника НАДФ*Н2 для синтеза жирных кислот:
а) ГМФ-путь распада углеводов
б) В реакции окисления малата (смотрите рисунок).

Эта реакция протекает в цитоплазме и катализируется ферментом МАЛАТДЕГИДРОГЕНАЗОЙ ДЕКАРБОКСИЛИРУЮЩЕЙ, тривиальное название которого – ЯБЛОЧНЫЙ ФЕРМЕНТ.
4. В ходе синтеза жирных кислот в каждом цикле удлинения используется не сам Ацетил-КоА, а его производное - малонил-КоА (при b-окислении каждый цикл укорочения приводит к образованию Ацетил-КоА).

Эту реакцию катализирует фермент АЦЕТИЛ-КоА-КАРБОКСИЛАЗА. Это ключевой фермент в мультиферментной системе синтеза ЖК. Этот фермент регулируется по типу отрицательной обратной связи. Ингибитором является продукт синтеза: ацил-КоА с длинной цепью (n=16) - пальмитоил-КоА. Активатором является цитрат. В состав небелковой части этого фермента входит витамин H (биотин).
Далее происходит поэтапное удлинение молекулы Ац-КоА на 2 углеродных атома за каждый этап за счет малонил-КоА. В процессе удлинения малонил-КоА теряет СО2. После образования малонил-КоА основные реакции синтеза жирных кислот катализируются одним ферментом - синтетазой жирных кислот (фиксирован на мембранах эндоплазматического ретикулума). Синтетаза жирных кислот содержит 7 активных центров. Участок, связывающий малонил-КоА, содержит небелковый компонент – витамин B3 (пантотеновую кислоту).
|
|
|
