 |


Реакции синтеза жирных кислот.
|
|
|
|
РЕАКЦИИ СИНТЕЗА ЖИРНЫХ КИСЛОТ.



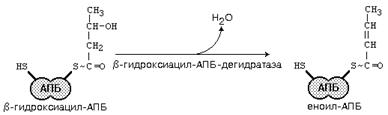

После этого ацил-АПБ вступает в новый цикл синтеза. К свободной SH-группе ацилпереносящего белка присоединяется новая молекула малонил-КоА. Затем происходит отщепление ацильного остатка, и он переносится на малонильный остаток с одновременным декарбоксилированием, и цикл реакций повторяется.
Таким образом, углеводородная цепочка будущей жирной кислоты постепенно растет (за каждый цикл – на два углеродных атома). Это происходит до момента, пока она не удлинится до 16 углеродных атомов (в случае синтеза пальмитиновой кислоты) или более (синтез других жирных кислот). Вслед за этим происходит тиолиз, и образуется в готовом виде активная форма жирной кислоты – ацил-КоА.
УСЛОВИЯ ДЛЯ ПРОТЕКАНИЯ СИНТЕЗА ВЫСШИХ ЖИРНЫХ КИСЛОТ:
1) Поступление углеводов, при окислении которых образуются необходимые субстраты и НАДФН2.
2) Высокий энергетический заряд клетки – высокое содержание АТФ, которое обеспечивает выход цитрата из митохондрий в цитоплазму.
Обмен углеводов и обмен жиров очень тесно связаны. Углеводы легко могут превращаться в жиры, а вот превращение жиров в углеводы невозможно. Жиры не могут превращаться в углеводы, так как Ацетил-КоА не может превращаться в пируват. Обмен жиров и углеводов объединяется как энергетический обмен, который находится под контролем гормонов.
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ СИНТЕЗА ЖИРА
Основным гормоном, регулирующим липогенез, является инсулин. Инсулин стимулирует синтез жира. На генетическом уровне инсулин стимулирует биосинтез ферментов, катализирующих образование ацил-КоА и триацилглицеринов. Инсулин также стимулирует биосинтез ферментов, обслуживающих обмен липидов – ферментов ГМФ-пути распада углеводов и яблочного фермента. Поэтому истощенным больным вводят глюкозу одновременно с инсулином с целью увеличения жировых запасов.
|
|
|
ТРАНСПОРТ ЭНДОГЕННОГО ЖИРА И ХОЛЕСТЕРИНА ИЗ ПЕЧЕНИ В ДРУГИЕ ТКАНИ.
Транспортной формой эндогенного жира и холестерина из печени в другие ткани являются ЛОНП - липопротеины очень низкой плотности. Ядро этих частиц состоит из эндогенных триацилглицеринов и холестерина, а оболочка - из фосфолипидов, белка апоВ100 (образуется в печени, молекулярная масса 100кДа).
Из печени ЛОНП поступают в кровь, где к ним присоединяются белки апоЕ и апоС.
АпоС - активатор липопротеин-липазы капилляров, расщепляет триглицериды, образуются глицерин и жирные кислоты - они поступают в ткань. Затем ЛОНП теряет апоС, превращаясь в ЛПП - липопротеины промежуточной плотности. ЛПП, теряя апоЕ, превращается липопротеины низкой плотности (ЛНП). Он содержит мало триглицеридов и много холестерина. Его функцией является перенос холестерина из печени в ткани.

Рецепторы к В100 были открыты американскими биохимиками Гольдштейном и Брауном. При генетическом дефекте рецепторов к B100 развивается наследственная гиперхолестеринемия, приводящая к раннему атеросклерозу. В этом случае у гомозиготных детей уже в возрасте 5-7 лет наблюдаются множественные инфаркты миокарда. У гетерозигот острые инфаркты миокарда отмечаются в 30-40-летнем возрасте. Если изменяется конформация В100, нарушается взаимодействие лиганда и рецептора. Фактор риска для этой модификации – воздействие на ЛНП компонентов табачного дыма.
|
|
|
