 |


Мобильные генетические элементы.
|
|
|
|
МГЭ - участки генет-й цепи материала (фрагменты ДНК или сегменты хромосом), кот. способны перемещаться и менять свое местопол-е в геноме.
Процесс перем-я МГЭ – транспозиция. МГЭ были открыты в 1947 г. Барбарой МакКлинтон на кукурузе. Для того чтобы объяснить появл на зернах кукурузы пигментированных пятен различ-х форм и размеров, она высказывала предположение о сущ-нии МГЭ, кот отвечают за появление этих пятен. Подтверждением данному предполож-ю послужили набл-я разрыва хро-м, проис-х в кл-х исслед-х раст-й кукурузы.
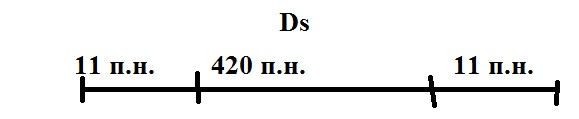
По Б. МакКлинтон у кукурузы сущ-ет 2 МГЭ, контр-их обр-е пигментных пятен: Ac – активатор, Ds – диссациатор.
Схема работы сис-мы Ac и Ds у кукурузы


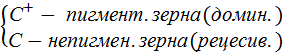
У некот. линий кук-зы неокр. зерна развив. в рез-те мутации С+→С обусл. встраивание в ген С+ МГЭ Ds.
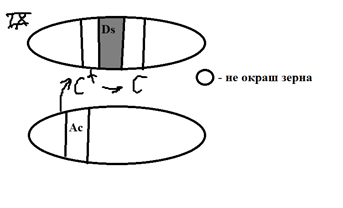 На другой хро-ме кук-зы расп. МГЭ – Ac, кот. отвеч. за транспозицию эл-та Ds. Т.е. контролир вырезание Ds и встраивание его в др. уч-ки хр-м.
На другой хро-ме кук-зы расп. МГЭ – Ac, кот. отвеч. за транспозицию эл-та Ds. Т.е. контролир вырезание Ds и встраивание его в др. уч-ки хр-м.
Размерн. зависит от времени транспозиции: чем раньше произошла – тем больше пятна. С развития науки начались накоп. Данные о сущ-и МГЭ у др-х объектов: E. Coli (в 60-х г.), дрозофилы(1976 Гершев, Гвоздев).
В 1983 МакКлинтон удостоена нобелевскую премию по медицине за открытие МГЭ. В 1984 Нина Федоров при помощи молек. Методов выделена МГЭ, Ac и Ds и изучена их молек. строение.
Молекулярное строение Ac и Ds
Ас:
Инвентиров. повторы (полиндромы)
Ac – отвечает за синтез фер-та транспозаза, кот. осущ. вырезание или встраивание его в др. уч-ки хр-м.
 Ds – обр-ся в рез-те делеции уч-ка центр. сис-мы Ac.
Ds – обр-ся в рез-те делеции уч-ка центр. сис-мы Ac.

Благодаря инвертируемым повторам Ds может обр. стебель петлю, кот. узн-ся фер-ом транспозазой и замык-ся «ключ-замок», «рука-перчатка» и вырез. Из хр-мы.
|
|
|
МГЭ прокариот
Началу изуч. МГЭ положило открытие в конце 60-х г., необычных мутантов по, общими для этих мутантов были вставки стр-ах, кот приводили к мутациям и были названы Ys – элем-ты.
YS – элем-ты обл-ют способ-ю менять свое местоположение на хр-ме, т.е. явл. МГЭ.
Им-ся 2 разнов-ти:
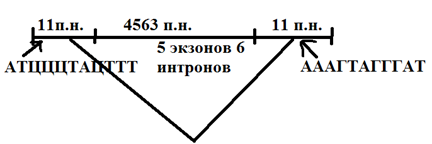
1) Ys-элем-ты - их размеры =200-5700 п.н. он хар-ся след особенностями:
· На концах несут несовершенные повторы (палиндромы), длиной от нескольких пар до неск. десятков пар нуклеотидов.
· Содержат ген кодирующий синтез транспозаза, отвеч. за перемещ. Ys-ов.
· При встраивании в др. уч-ки хр-м небольшие дупликации длина 5-11 п.н. (флакирующие Ys-эл-ты).
Бакт. Хр. E.Coli несет только Ys-ов в кл перемещ с частотой 10-6-10-8 на клет деление.
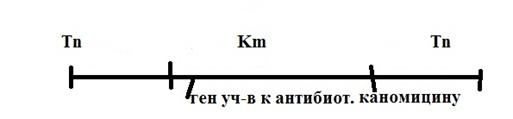
2) Tn – элементы (транспозоны) харак. теми же особ., что и Ys кроме гена кодир. транпозазу, они могут переносить гены устойч-ые к антибиотикам. Эти гены могут расп-ся как внутри Tn так и между 2-мя Tn-эле-ми.
МГЭ эукариот
Транспозоны
Пример: - Ac, Ds у кукурузы,
-P- эле-ы у дрозофилы
По краям не несут инвентир. повторы, а в центр. част. ген, кодир. синтез фер-та транспозаза.
В 1976 г. Гвоздев, Георгиев открыли МГЭ у дрозофилы, кот получили назв Мобильные Дисперсированные Гены (МДГ).
МДГ случайно распред. по всему геному и способ. переем-ся с частотой 1*10-5 – 10-8 на поколение.
В наст. Время у дрозофилы открыто множество МГЭ кот. Названы терминами обозн. Склонности к переем.:
Hobo – бродяга,
flea – блоха,
магелан
бигль
Ретротранспозоны
Получ. Такое название потому что в их перемещ приним. участие обратная транскрипция (ретро-обратный). Широко распространены у млекоп. сост. более 10% от всей ДНК
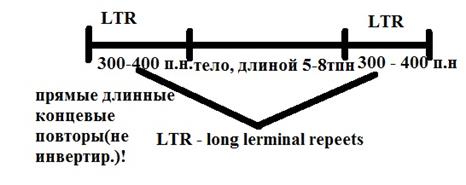
Механизм перем-я ретротранспозонов

Значение МГЭ:
1. Увелич. Генет. измен-ти перемещ. по геному и встраив. в гены приводят к увелич. частоты мутаций.
2. Измен-ие экспрессии генов – встраива-ет гены МГЭ меняя хар-р их экспрессии (могут полностью выключать гены или измен. интен-ть их работы).
|
|
|
3. Исп-ся в качестве векторов в генной инженерии.
ГЕНЕТИЧЕСКАЯ РЕКОМБИНАЦИЯ
Г.Р. - совок. процессов приводящ. к перераспред. нукл. послед-й и в геномах. Явление г.р. было открыто в 1910 г. Т. Морганом на дрозофиле. В частности им был установлен обмен уч-ми между гомолог. хр-ми получ. назв. – кроссинговер ( один из частных случаев рекомбинации).
К Г.Р. след. процессы:
-обмен уч-ми между молек. ДНК (или РНК)
-встраивание уч-в ДНК в опред. места на молек-х ДНК.
-комбинирование хр-м при формировании гамет и спор
-кроссинговер – обмен уч-ми между гомолог. хр-ми
В настоящее время Г.Р. подразд на 2 класса:
1) общая
2) сайт-специфическая(«незаконная»)
1. Общая: протекает на основе гомологии рекомбин. уч-в ДНК
2. «незаконная»: для ее осущ. гомологии реком. уч-в
Общая рекомбинация
Наиболее изученным процессом отн. к общей рекомб. явл. кроссинговер, кот. происх. в профазе мейоза1 у эу-.
На этой стадии хр-м сост из 2-х хроматид в обмене уч-т 2 хроматиды из 4, каждая из хроматид содер. 1 мол. ДНК.

|
|
|
