 |


2.Lack of visualization of the fetal nasal bone
|
|
|
|
2. Lack of visualization of the fetal nasal bone
At the end of the 1st trimester nasal bone is not determined by ultrasound at 60-70% of fetuses with Down syndrome and in 1-2% of healthy fetuses. Agenesis or hypoplasia of the nasal bone in fetuses with Down syndrome occurs due to delay ripening centers of ossification of the nasal bones.
ADDITIONAL NON-INVASIVE METHODS OF PRENATAL DIAGNOSIS
The fetal echocardiography plays more and more important role in prenatal diagnosis of congenital fetal diseases. At first, patients receive timely information about the prognosis of the disease of children, about treatment options, and directed to provide skilled assistance in special hospital. Secondly, the detection of congenital heart disease initiates a research aimed at the identification of hereditary diseases and syndromes. Third, the studies help to resolve the issue of prolongation of pregnancy or medical interrupt of it. Transvaginal fetal echocardiography is an expert study, should be conducted at the PD centers.
The main indications for fetal echocardiography are:
· positive family history (congenital heart disease in the parents or earlier born children);
· abnormal image of the heart and/or major arteries when screening ultrasound;
· extracardiac anomalies;
· NFT expansion zone.
To assess uterine blood flow Doppler ultrasound is used. Doppler ultrasound evaluate the blood flow in the aorta and internal carotid artery of the fetus, in umbilical vein and arteries, in the uterine and ovarian arteries.
Magnetic resonance imaging (MRI) - is a highly informative method of prenatal diagnosis and can be recommended in various pathological processes and fetal anomalies, suspected by ultrasound, especially in cases of central nervous system malformations.
A computer program PRISCA
PRISCA - a computer program for calculating the risk of congenital abnormalities of the fetus, taking into account the data of physical gynecological examination, ultrasound examination and serum marker data, as well as the influence of other factors.
PRISCA allows:
1. to calculate the probability of different types of fetal abnormalities:
-trisomy 21 (Down syndrome)
-trisomy 18 (Edwards syndrome)
-CNS anomalies (neural tube defects);
2. to take into account the individual data of a patient;
3. to take into account the factors influencing the detection of deviations from normal levels of biochemical markers.
The calculation takes into account factors:
· maternal age
· term of pregnancy
· body weight
· ethnicity
· smoking
· Diabetes mellitus
· the number of fetuses in the uterus
· in vitro fertilization.
The calculation in the 1st trimester
The recommended terms of testing: 9-14 week of pregnancy according to the LNMP.
The calculation takes into account following factors:
· early term of pregnancy
|
|
|
· high level of pathology on condition of PAPP-A + free β -CHG+ NT
· detection sensitivity – 90%
· false-positive results – 5%
Output: age risk, biochemical risk, calculation of risk of trisomy 18 and trisomy 21 in PAPP-A + free β -CHG+ NT. considering the age of the pregnant
The calculation in the 2nd trimester
The recommended terms of testing: 14 – 21 week of pregnancy (the most optimal in 16-20 weeks).
The calculation takes into account following factors:
· US (term of pregnancy
· serum markers ( AFP, HCG, uE3).
Output: age risk, biochemical risk, calculation of risk of trisomy 18 and trisomy 21 and neural tube defects by AFP=HCG+ uE3+ NT, considering the age of the pregnant.
Peculiarities of risk calculation;
1. The calculation of risk depends on the accuracy of the data, used for analysis.
2. The calculation of risk is a result of statistic analysis.
3. The results should be confirmed (or excluded) by cytogenetic studies.
4. The issued result indicates degree of risk for various pathologies of the fetus in the form of text and graphics, specify the values of specific markers.
New directions in prenatal diagnosis
One of the promising approaches in PD is the search for genetic markers.
Analysis of genes of the major histocompatibility complex of humans (HLA) is positively proved itself in the diagnosis of many human diseases, including in pediatric patients, particularly in children with cerebral palsy.
Molecular-genetic methods have been increasingly used for PD, and the most important of them is a cytogenetic technique of fluorescence in situ hybridization (FISH). It is based on the use in the reaction of in situ hybridization of various DNA samples.
Rapid and economical screening method for determining aneuploidy and fetal triploidy is a test based on quantitative fluorescent multiplex PCR (polymerase chain reaction).
At the present time in connection with the development of techniques of in vitro fertilization (IVF) methods of preimplantation genetic diagnosis (PGD) for chromosomal diseases becoming more widespread. The advantage of this approach is that it rules out the therapeutic abortion, since chromosomal and genetic disorders are defined on oocytes and embryos and this makes it possible to transfer into the uterine cavity intact embryos (Fig. 2).
PGD is performed for the diagnosis of hereditary diseases. PGD can be carried out in situations where there is an increased risk that the embryos will have certain chromosomal abnormalities. These abnormalities may reduce the probability of implanting (attaching) in the uterus, may lead to miscarriage or the birth of a child with physical or mental disabilities. PGD helps prevent this unfavorable outcome, because it allows to select healthy embryos before transfer into the uterus.
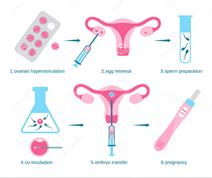
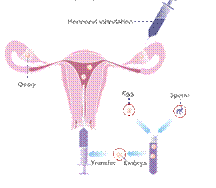
Fig. 2 The scheme of preimplantation genetic diagnosis
CHAPTER 5. Fertilization. The development of embryonic membranes and placenta
Fertilization
Fertilization is the fusion of a spermatozoon (a mature male germ) with an ovum (a female reproductive cell) into one cell which gives the start of a new organism.
The maturation of the ovum and spermatozoon is a complicated process during which the quantity of chromosomes in the nuclei of both cells is halved. The nucleus of a new cell resulting from fertilization contains a complete set of chromosomes.
|
|
|
Oogenesis
The process of development of a mature ovum is called oogenesis. The primitive germ cells take their origin from the yolk sac at about the end of the 3rd week and their migration to the developing gonadal ridge is completed round about the end of the 4th week. In the female gonads, the germ cells undergo a number of rapid mitotic divisions and differentiate into oogonia. The number of oogonia reaches its maximum by the 20th week, numbering about 7 million.
While the majority of the oogonia continue to divide, some enter into the prophase of the first meiotic division and are called primary oocytes. These are surrounded by flat cells and are called primordial follicles and are present in the cortex of the ovary. At birth there is no more mitotic division and all the oogonia are replaced by primary oocytes which have finished the prophase of the first meiotic division and remain in resting phase between prophase and metaphase. Total number of primary oocytes at birth is estimated to be about 2 million. The primary oocytes do not finish the first meiotic division until puberty is reached. At puberty about 400. 000 primary oocytes develop further, the rest being atretic. Out of these, some 400 are likely to ovulate during the entire reproductive period.
Maturation of the oocytes
The essence of maturation is reduction of the number of chromosomes to half. Before the onset of first meiotic division, the primary oocytes double its DNA by replication, so they contain double the amount of normal protein content. There are twenty-two pairs of autosomes which determine the body characteristics and one pair of sex chromosomes, named " XX". The first stage of maturation occurs with full maturation of the ovarian follicle just prior to ovulation but the final maturation occurs only after fertilization.
The primary oocyte undergoes the first meiotic division giving rise to secondary oocyte and one polar body. The two are of unequal size, the secondary oocyte contains haploid number of chromosomes (23, X), but nearly all the cytoplasm and the small polar body also contains half of the chromosomes (23, X) but with scanty cytoplasm. Ovulation occurs soon after the formation of the secondary oocyte.
The secondary oocyte completes the second meiotic division (homotypical) only after fertilization by the sperm in the Fallopian tube (Fig. 3) and results in the formation of two unequal daughter cells, each possessing 23 chromosomes (23, X), the larger one is called the mature ovum and the smaller one is the second polar body containing the same number of chromosomes. The first polar body may also undergo the second meiotic division. In the absence of fertilization the secondary oocyte does not complete the second meiotic division and degenerates as such.
Chromosome nomenclature. The number designates the total number of chromosomes (in numerals) followed by the sex chromosome constitution after the comma.

Fig. 3. Development of the ovum and its passage through the fallopian tube into the cavity of the uterus.
Structure of a mature ovum
A fully mature ovum is the largest cell in the body and is about 130 microns in diameter. It consists of cytoplasm and a nucleus with its nucleolus which is eccentric in position and contains 23 chromosomes (23, X). During fertilization the nucleus is converted into a female pronucleus. The ovum is surrounded by a cell membrane called vitelline membrane.
There is an outer transparent mucoprotein envelope, the zona pellucida. The zona pellucida penetrated by tiny channels which are thought to be important for transport of materials from the granulosa cells to the oocyte. In between the vitelline membrane and the zona pellucida there is narrow space called perivitelline space which accommodates the polar bodies. The human oocyte, after its escape from the follicle, retains a covering of granulosa cells known as the corona radiata derived from the cumulus oophorus.
|
|
|
Spermatogenesis
The process involved in the development of spermatids from the primordial male germ cells and their differentiation into spermatozoa is called spermatogenesis. Shortly before puberty, the primordial germ cells develop into spermatogonia and remain in the wall of seminiferous tubules. The spermatogonia, in turn, differentiate into primary spermatocytes which remain in the stage of prophase of the first meiotic division for a long time (about 16 days). Each spermatocyte contains 22 pairs of autosomes and one pair of sex chromosomes named " XY". With the completion of the first meiotic division, two secondary spermatocytes are formed having equal share of cytoplasm and haploid number of chromosomes either 23, X or 23, Y. This is immediately followed by the second meiotic divisioin (homotypical) with the formation of four spermatids, each containing haploid number of chromosomes, two with 23, X and two with 23, Y. Immediately after their formation an extensive morphological differentiation of the spermatids occurs without further cell division to convert them into spermatozoa. The process is called spermiogenesis. In man the time required for a spermatogonium to develop into a mature spermatozoon is about 61 days.
The spermatozoa are formed in contorted seminiferous tubules of the male sex glands. A spermatozoon comprises a head, a neck, and a long thin tail. The head is the nucleus of the germ surrounded by a thin layer of cytoplasm, while the neck and the tail are cytoplasm proper.
The motility of a spermatozoon depends on the movements of its tail. The spermatozoon acquires the power of motility after it is mixed with the prostatic fluid and the secretion from the seminal vesicles. The mixture of spermatozoa and the secretion from the seminal vesicles and the prostate is the sperm or semen.
About 5-8 ml of the seminal fluid are ejaculated into the vagina during coitus. This portion contains 200-500 million spermatozoa. The semen is mainly accumulated in the posterior fornix into which the cervical portion of the uterus is projected. The external os of the endocervical canal is bathed in the sperm accumulated in the posterior fornix which provides favourable conditions for fertilization.
During sexual excitement the uterine muscles are contracted to open the external os of the cervix slightly. The mucous stopper of the cervix is protruded into the semen deposited in the posterior fornix during copulation. After coitus the mucous stopper is pulled back into the cervix together with the spermatozoa.
The main condition for penetration of the spermatozoa into the uterus is however their own motility. The vagina has an acid medium and the spermatozoa move toward the uterine cervix where the reaction is alkaline and favourable to their reproductive function. The alkalinity of the uterus and the fallopian tubes maintains the reproductive potential of spermatozoa and their motility for a few days. At ovulation the cervical mucus becomes less viscid, facilitating rapid transit of sperm from the vagina to the endometrial cavity. Sperm may live in the vagina for up to 3 days before ovulation. They can migrate to the fimbriated end of the uterine tube in 5 min under experimental conditions.
Spermatozoa travel at a rate of 3-3. 5 mm per min. In 60-90 minutes they can be found in the uterus cavity, and in two or three hours in the fallopian tubes where they meet the ovum.
|
|
|
Fertilization usually occurs at the ampullar portion of the tubes.
The ovum cannot move by itself. The peristaltic activity of the tube and the motion of its ciliated lining induce a current of fluid which entraps the ovum and propels it from the infundibulum to the uterine end of the tube. Sperm and ovum unite resulting in a zygote which continues dividing and developing during its transit through the tube to the endometrial cavity. One of spermatozoa enters the ovum through tubercle on its surface and get into the protoplasm of the ovum. This penetration is facilitated by hyaluronidase, the enzyme produced by the spermatozoon, which dissolves the transparent membrane of the ovum, mucinase, the trypsin-like enzyme, etc. The nucleus of spermatozoon moves toward the nucleus of the ovum and the fusion of the nuclei is followed immediately by the division process. The zygote makes its journey through the tube and moves to the site of implantation in 8–10 days. During this time the conceptus is dividing; at the time of implantation it has formed a blastocyst (a single layer of cells surrounding a central cavity). One area of the blastocyst wall is 3 to 4 cells thick. This area is the embryonic pole, which soon becomes recognizable as an embryo.
Implantation
Qualitative changes also take place in a fertilized ovum along with the mitosis. When in the fallopian tube, the fertilized egg is separated from its transparent coat (zona pellucida). By the moment the ovum enters the uterus, the outer layer of the morula cells is converted into the trophoblast, and the inner cells — into the embryoblast. The embryoblast develops into the embryo.
The trophoblast is a layer of tissue which attaches the ovum to the uterine wall and supplies nutrition to the embryo developing from the embryoblast.
The cells of trophoblast secrete enzyme-like substances which dissolve the tissues of the uterine mucosa. As the ovum sets on the superficial layer of the endometrium (normally on the anterior or posterior wall, at the level of insertion of the tubes), its trophoblast dissolves the epithelial lining, the glands, the stromal cells and the vessels of the uterine mucosa and is gradually embedded into the functional layer of the endometrium. When the ovum is completely embedded into the mucosa, the site of entrance becomes sealed. This moment is assumed to mark the completion of the implantation process. The human blastocyst implants itself on the wall of the uterus, probably during the late sixth or early seventh day of its development, on endometrium that may range from the eighteenth to the twenty-third day of its development.
The endometrium is at this moment in its secretory phase and contains substances necessary for the nutrition of the embryo. (Fig. 4)
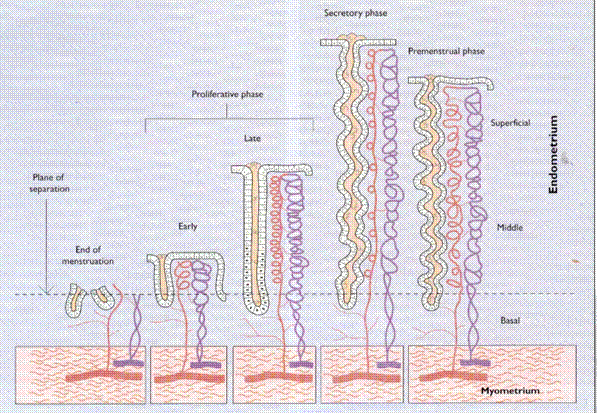
Fig. 4. Endometrial vascular pattern.
When the uterine mucosa is dissolved by the trophoblast, the tissues around the settled ovum disintegrate to give nutrition to the embryo (embryotroph). The embryotroph contains proteins, carbohydrates, fats, vitamins, salts and other substances necessary for the nutrition and growth of the embryo.
The trophoblast proliferates at the site of implantation, which at first consists of solid cytotrophoblast and syncytiotrophoblast. The latter becomes vacuolated on the eighth day to develop lacunae for the reception of maternal blood on about the eleventh day. The chorionic villi begin to form as cytotrophoblastic masses on the twelfth to the thirteenth day and grow peripherally along the syncytiotrophoblastic framework, ultimately coalescing peripherally and displacing the syncytiotrophoblast, except the portion lining the intervillous space. Remnants of the desquamated syncytiotrophoblast are encountered in the placental site as giant cells.
Development of the fetus and fetal membranes
Following the implantation, the ovum rapidly grows. Projections (villi) are formed on the trophoblast. At this stage they have no blood vessels and are actually primordial villi. The outer coat of the ovum is called the chorion. Tissues between the villi and the endometrium disintegrate and the maternal blood starts circulating. The space between the chorionic villi and the surrounding endometrium is the primary intervillous space, which surrounds the entire ovum. At later terms the intervillous space remains only in the region of the placenta and is called the secondary intervillous space.
Simultaneously the inner cells of the ovum, embryoblast, develop. The development of the embryoblast begins when the ovum is in the tube. Cavities filled with fluid are formed in the middle of the embryoblast, which are then fused into one cavity, exocoelom, and the ovum transforms into the blastocyst. The cells surrounding the blastocyst cavity form the mesoblast.
|
|
|
Cells accumulate in one segment of the blastocyst. Two nodules can be distinguished in this accumulation, the ectoblast and the entoblast. Cavities are soon formed in the centres of these nodules and the ectoblast turns into the ectoblastic vesicle. The ectoblastic vesicle is connected through a stalk with the trophoblast which then forms the amniotic cavity. The wall of this cavity turns into a fluid sac, the amnion. The entoblastic vesicle is situated closer to the centre. Later it develops into the yolk sac.
The ectoblast and entoblast cells situated between the amnion and yolk sac form the embryo.
The size of the exocoelom cavity increases and the mesenchimal cells are displaced toward the chorion (trophoblast), amnion, yolk sac, and the embryo in between them. The walls of the vesicles and the chorion become double layered. The embryo is composed of three layers: the ectoderm, mesoderm and entoderm. All tissues and organs of the fetus develop from these three layers.
The amnion rapidly increases in size due to the accumulation of fluid in the sac, while its wall (the amnion proper) moves toward the villous membrane and becomes attached to it. The blastocyst cavity disappears. The embryo, which was situated between the amnion and the yolk sac, starts screwing into the amnion cavity and is gradually embedded into it. As the amniotic cavity increases in size, the yolk sac decreases and disintegrates.
At the same time allantois starts to form from the hindgut of the embryo. The allantois extends to the villous membrane along the stalk that connects the amniotic vesicle with the trophoblast. The allantois envelops vessels that course from the embryo body to the villous chorion. The vessels grow into each villus of the chorion. At this stage the allantoic blood circulation of the embryo is started.
Soon after that, the fetus becomes surrounded by the amniotic fluid and three membranes: decidual membrane, the chorion, and the amnion. The chorion and the amnion belong to the fetus, while the decidual membrane is maternal (it is formed from the functional layer of endometrium).
The layer of mucous membrane which covers the implanted ovum is called the decidua capsularis; that portion on which the ovum rests, the decidua basalis; all the rest of the lining of the uterus, the decidua vera.
Microscopically, examination of an early decidua vera shows: (1) the epithelium, best preserved around the openings and deeper portions of the glands; (2) the compact layer of the decidua, nearly one half of the whole thickness, made up of the swollen and hyperplastic fibrillary stroma and decidual cells which are large, ovoid or spindle-shaped, with much protoplasm, and with single or rarely several large vesicular nuclei, and are derived from the stroma; (3) the glandular or ampullary layer, or spongiosa, consisting of the enlarged, elongated and convoluted uterine glands, often with papillary growths of the epithelium which is low and cuboid without cilia and contains fewer decidual cells; and (4) a small, unaltered layer through which the fundi of the glands pass to, and sometimes slightly into the muscular coat. The glands pursue a straighter course in the basal portions of the compacta than in the spongiosa. Around the insertion of the ovum the blood vessels are most numerous and the capillaries most dilated. The veins are enlarged and open into the intervillous spaces. The glands are pushed aside by the growing ovum.
The decidua basalis is a portion of the decidua vera altered only by the superimposed growing ovum. It has the compacta and spongiosa, but the glands and blood vessels, the latter enormously developed, are stretched to a plane parallel to the wall of the uterus and at the junction of the capsularis curve in arches upward into it. The portion of the basalis next to the chorion or trophoblast is called the trophosphere.
The decidua capsularis is that portion of the vera which is stretched over the ovum. It is thick at the equatorial portion and thin at the outer pole. It is at this point that the egg buries itself in the mucosa. This point, which is microscopic, is soon covered over. The portions near the base resemble the vera; the blood vessels are more numerous; the veins are larger, and the capillaries open into intervillous spaces. The glands are arranged tangentially around the ovum, and one finds glands opening obliquely on the surface (not into the cavity of the trophosphere) almost up to the vertex of the capsularis.
As the ovum grows and fills out the cavity of the uterus, the decidua undergoes many changes. The capsularis at first is stretched over the ovum, and soon reaches the opposing wall of the uterus, on which it lies, but does not adhere. The uterine cavity is completely obliterated by the fourth month. The capsularis is fused with the vera and the chorion and regresses but is readily recognized at the internal os. The chorion adheres to the vera and on the delivered secundines large portions of the vera are found attached to the chorion. At the edges of the placenta one usually finds a thick layer of chorionic ectoderm in fibrinous degeneration, the site of the transition of vera and basalis into capsularis.
The decidua vera grows rapidly in thickness until the beginning of the third month; from then on it thins out until at term its thickness is only 1 mm or less. The epithelium, except in the glands of the lowest layers of the spongiosa and those between the muscle bundles, has disappeared. From these remnants the new uterine mucosa is reconstructed. The compacta is reduced to a narrow strip, and most of the decidual and stroma cells are in process of coagulation necrosis. The glands of the compacta are separated from each other, while in the spongiosa they form a thin, loose, flattened network, and since the stroma here is also beginning to degenerate, separation of the ovum from the uterine wall is easy. The basalis undergoes the most decided transformation, because here the egg is inserted and the placenta is situated.
Placentation
The first evidence of placental formation is the development of trophoblast cells by day 10. Invasion of these cells into maternal blood vessels causes blood to leak into the space between cells, forming lacunae (lakes), which become the intervillous space. The fetus derives nourishment from the lacunae. Initially, the placenta surrounds the blastocyst, transmitting nutrients and discharging wastes directly across cell membranes. As early as day 11 or 12, villi begin to form on the chorionic surface; they branch and rebranch in a complicated treelike arrangement around the chorionic surface. Villous transfer from maternal blood to fetal blood begins when fetal vessels appear within the placenta by day 19.
At about 12 weeks, apparently influenced by the location of the major source of the maternal blood supply, the true (discoid) placenta begins to demarcate at the old embryonic pole; the placenta is attached by villi to the decidua directly overlying maternal spiral arterioles. The other villi atrophy, disappearing entirely by 16 to 18 week. The spiral arterioles empty into the intervillous space so that maternal blood circulates around and through the latticework of villi, draining out through two or three venous sinuses associated with each spiral arteriole. The villi are divided into groups called cotyledons, each supplied by one or two spiral arterioles. At term, a placenta contains 10 to 20 cotyledons. Nutrients are transferred from maternal blood in the intervillous space, across trophoblast cells, through the fibrous core of the villus, and through the endothelial cells of the fetal capillaries to the fetal blood. Wastes move in the opposite direction. This arrangement is called a hemochorial placenta, because maternal blood is in apposition to fetal chorionic or trophoblastic tissue. (Fig 5)
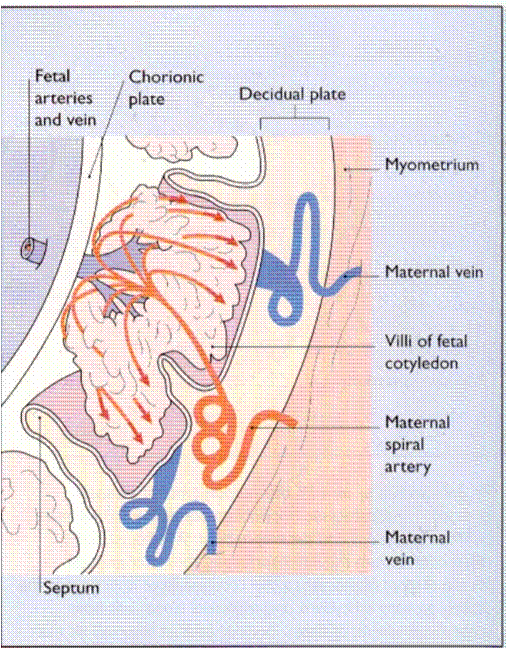
Fig. 5. The diagram of human placenta
The discoid placenta reaches its final form at 18 to 20 week of pregnancy. It continues to grow throughout pregnancy, weighing about 600 g at delivery.
Placenta produces a complex of hormones in a maternal organism, such as:
• Chorionic gonadotropin, which is instrumental in lengthening of function of yellow body in the first weeks of pregnancy; together with prolactin, it facilitates transformation of yellow body of menstruation in to the yellow body of pregnancy; in the early terms of pregnancy chorionic gonadotropin stimulates steroidogenesis in the yellow body of ovary. In the second half of pregnancy chorionic gonadotropin stimulates the synthesis of estrogens in a placenta, participating in aromatization of androgens; chorionic gonadotropin takes part in differentiation of the fetal sex.
• Estrogens, which are produced by chorion in quantities greater than in ovaries. Estrogens affect metabolic processes and growth of uterus causing hyperplasia and hypertrophy of endometrium and miometrium, taking an active part in development of labor process.
• Progesteron which appears from maternal cholesterol in chorion. Its biological role in development of pregnancy is great: it participates in implantation of fertilized ovum, suppresses uterine contractility and supports the tone of isthmicocervical part of the uterus, stimulates the growth of uterus at pregnancy, participates in steroidogenesis. It also renders an immunosuppressive action, that is important for development of embryo.
• Placental lactogen which is synthesized by the cells of trophoblast, matters very much in development of pregnancy, because possesses prolactin activity and immunological properties of growth hormone. The basic biological role of placental lactogen consists in adjusting of carbohydrate and lipidic metabolism, strengthening of synthesis of albumen in the organism of fetus, resulting in the mass of fetal body.
• Prolactin produced by chorionic tissue and decidual membrane performs a definite role in the production of pulmonary surfactant of the fetus and adjusting of fetoplacental osmoregulation.
Thus, the hormone-producing function of placenta determines the basic physiological processes in the system “mother — placenta — fetus”.
Functions of the placenta
• Respiratory function of placenta: placenta provides the delivery of oxygen from mother to the fetus, and taking away of carbon dioxide from the fetus to mother.
• Nutrition of the fetus and excretion of assimilation products. Transport of albumens, fats, carbohydrates from the blood of mother into the blood of fetus is realized due to difficult processes of the enzymatic conversion.
The syncytiotrophoblast of the placenta produces specific proteins and glycoproteins, which participate in the processes of deaminization of amino acids, their synthesis from predecessors, are instrumental in the active transport of them to the fetus. Lipids of placenta are synthesized in a placenta from predecessors being found in the blood of mother. 2/3 lipids of placenta make up phospholipids, which take part in biosynthesis of albumen, adenosine triphosphate (ATP), DNA, in the transport of electrolytes and amino acids, in adjusting of fermentative activity of placenta. Phospholipids provide permeability of membranes of placental cells for different substances.
• Immunologic defence of the fetus: placenta is an immune barrier between two genetically foreign organisms (mother and fetus), preventing the immune conflict.
• Protective function of placenta: placenta protects an organism from unfavourable influence of harmful factors getting in the organism of mother (toxic substances, medicinal preparations, microorganisms). However, a barrier function of placenta is selective and for some damaging matters it turns out insufficient.
Self Test
1. Germ cells which provide the continuity of generations occur in the
A. gonads.
B. genital ridges.
C. yolk sac.
D. coelomic epithelium.
E. mesonephrons.
2. During development of the ovary the primordial germ cells migrate to the
A. wolffian ducts.
B. genital ridges.
C. dorsal mesentery.
D. mesonephric tubules.
E. urogenital diaphragm.
3. In the third month of a normal human pregnancy the ovary has approximately
A. ten million germ cells.
B. one million germ cells.
C. one hundred thousand germ cells.
D. one thousand germ cells.
E. one hundred germ cells.
4. Fertilization usually occurs at the
A. ampullar portion of the tube.
B. ovarian surface.
C. uterine cavity.
D. isthmus of the tube.
5. During normal human pregnancy the following takes place in the ovary:
A. ripening of the follicle
B. ovulation
C. absence of ovulation and maturation of follicle
D. a yellow body ripens and follicles do not ripen
6. The yellow body of pregnancy starts to be subjected to involution
A. during the 1st month of pregnancy.
B. by the 3-4th month of pregnancy.
C. during the 2nd month of pregnancy.
D. by the 8-9th month of pregnancy.
E. at term of delivery.
7. Chorionic gonadotropin is produced in
A. hypophysis.
B. ovaries.
C. placenta.
D. hypothalamus.
8. The endometrium at the moment of implantation is in its
A. secretory phase.
B. proliferative phase.
C. regenerative phase.
D. desquamation phase.
9. The following hormones are synthesized by the cells of trophoblast, apart from:
A. placental lactogen.
B. prolactin.
C. estrogens.
D. progesterone.
E. insulin.
10. The following structures cover to fetal membranes, except for:
A. amnion.
B. chorion.
C. decidual membrane.
D. albugineus.
CHAPTER 6. ANATOMY OF FEMALE REPRODUCTIVE ORGANS
The female reproductive organs are those which are concerned with copulation, fertilization, growth and development of the fetus and its subsequent exit to the outer world. The organs are commonly divided into:
• external genitalia
• internal genitalia,
• accessory reproductive organs.
External Genitalia
The external genitalia are visible at a visual inspection and are subdivided into mons pubis, labia majora, labia minora, clitoris, vestibule, and perineum. (Fig. 6).

Fig. 6 External genitalia. Mons pubis and labia majora.
Mons pubis. It is the pad of subcutaneous adipose connective tissue lying in front of the pubis and covered in the adult female by hairs. The upper bound of hair coverage in women is horizontal, in men — vertical.
Labia majora. The vulva is bounded on each side by the elevation of skin and subcutaneous tissue which form the labia majora. The labia majora are covered with squamous epithelium and contain sebaceous glands, sweat glands and hair follicles. Beneath the skin there is a dense connective tissue and adipose tissue. The adipose tissue is richly supplied. There are radiated terminations of the round ligaments of uterus here. Labia majora are often lacerated during childbirth with development of hematoma.
Labia minora. They are two thin folds of skin devoid of fat on either side just within the labia majora. Anteriorly they are divided into two ends: anterior and posterior. Anterior ends of both labia unite with each other above the clitoris forming its prepuce (praeputium clitoridis). Posterior ends unite with each other below the clitoris forming frenulum clitoridis. The lower portion of the labia minora fuses across the midline to form a fold of skin known as fourchette. It is usually lacerated during childbirth. The folds of labia minora contain connective tissue, numerous sebaceous glands, erectile muscle fibres and numerous vessels and nerve endings. (Fig. 7).

Fig. 7. External genitalia. Clitoris and labia minora.
Clitoris. It is a small cylindrical erectile body situated in the most anterior part of the vulva. It consists of the glans, body and two crura.
The glans is covered by squamous epithelium and is richly supplied with nerves and vessels. The vessels of the clitoris are connected with the vestibular bulb and are liable to injuries during childbirth.
Vestibule. It is a triangular space bounded anteriorly by the clitoris, posteriorly by the fourchette and on either side by the labia minora. The bottom of the vestibule is the hymen.
There are four openings into the vestibule:
• Urethral opening, which is situated in the midline just in front of the vaginal orifice about 1-1. 5 cm below the pubic arch;
• The paraurethral ducts open either on the posterior wall of the urethral orifice or directly into the vestibule;
• Vaginal orifice and hymen. The vaginal orifice lies at the posterior end of the vestibule. In virgins and nulliparae the opening is closed by the labia minora but in parous it may be exposed. Hymen is a septum of mucous membrane which is usually ruptured during the first coitus. During childbirth the hymen is extremely lacerated and is later represented as carunculae myrtiformes.
• Opening of Bartholin’s ducts. There are two Bartholin’s glands, one on each side. They are situated in the superficial perineal pouch, close to the posterior end of the vestibular bulb. They are pea-sized and yellowish-white in colour. Each gland has got a duct which measures about 2 cm and opens into the vestibule outside the hymen at the junction of the anterior two thirds and posterior one third in the groove between the hymen and the labium minus. Due to the block of the ducts bartholinitis may occur with the development of abscess.
Perineum. It is usually divided into anatomical and obstetrical perineum.
Anatomical perineum is bounded above by the inferior surface of the pelvic floor, below — by the skin between the buttocks and thighs. Laterally, it is bounded by the ischiopubic rami, ischial tuberosities and sacrotuberous ligaments, and posteriorly — by the coccyx. The perineum is divided into two triangular spaces: the anterior and posterior one. The anterior space is usually called the urogenital triangle; the posterior one is called the anal triangle.
Obstetrical perineum (central point of perineum). The pyramidal-shaped tissue where the pelvic floor, the perineal muscles and fascia meet in between the vagina and the anal canal is called the obstetrical perineum. It measures about 4 cm x 4 cm with the base covered by the perineal skin and the apex is pointed and is continuous with the rectovaginal septum.
Muscles and fascias of the perineum. The musculofascial structures include the following:
- Fasciae (two layers of perineal fascia: superficial and deep layers of perineal fascia, inferior and superior layers of urogenital diaphragm);
- Muscles: the pelvic floor is composed of three layers of muscles coated by fasciae: lower (outside), middle and upper (inside). (Fig. 8)
The lower (outer) layer. Four muscles make the lower (outer) layer: bulbocavernous muscle, ishiocavernous muscle, the external sphincter of anus, superficial transverse muscle of perineum. The muscles are arranged in a form of figure “8” suspended from the pelvic bones.
A musculofascial layer which surrounds urethra (anteriorly) and vagina (posteriorly) constitute the inner layer. This layer is called urogenital diaphragm. The muscles of urogenital diaphragm are deep transverse muscle of the perineum (m. transversus perinei profundus) and m. sphincter uretro-vaginalis. The muscle bundles of the anterior portion of this diaphragm surrounding the urethra and vagina form their sphincter and are called m. sphincter uretrovaginalis. The posterior portion of the urogenital diaphragm contains muscular bundles extending in the transverse direction to the ischial tuberosities (pair muscle). This portion of the urogenital diaphragm is the deep transverse muscle of the perineum (m. transversus perinei profundus).
The upper (internal) layer of perineum is called the pelvic diaphragm and consists of a pair muscles — m. levator ani.
Muscles and fascias of the perineum at the same time represent a supporting system of the uterus.
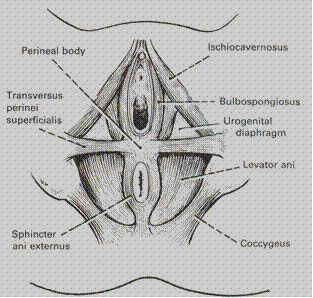
Fig. 8. Mucsles of the pelvic floor
Internal genitalia
The internal female genital organs include vagina, uterus, Fallopian tubes and ovaries. These organs are situated internally and cannot be visible, so they require special instruments for inspection.
Vagina. It is a fibromuscular-membranous sheath connecting the uterine cavity with the vulva. It constitutes the excretory canal for the uterine secretion and menstrual blood. It is the organ of copulation and forms the birth canal of parturition. The canal is directed upwards and backwards forming an angle of 45 degrees with the horizontal in erect posture. The long axis of the vagina lies almost parallel to the plane of pelvic inlet and at right angles to that of the uterus. The diameter of the canal is about 2. 5 cm being widest in the upper part and narrowest at its introitus. There is an anterior, posterior and two lateral walls of the vagina. (Fig. 9). The anterior and posterior walls are apposed together but the lateral walls are comparatively stiffer, especially in the middle, thus it looks “H”-shaped on transverse section. The length of the anterior wall is about 7 cm and that of the posterior wall is about 9 cm. The part of its cavity around the cervix is called fornix. Four fornices are distinguished: anterior, posterior and two lateral ones. The posterior fornix is much deeper and the anterior one is the most shallow.
In the adult nullipara longitudional furrows — the columns of the vagina — and numerous transverse ridges or rugae mark the vagina. This configuration permits great distension during parturition; at girls and women in postmenopauze the folds of vaginal wall are absent. The wall of vagina is 3 mm thick and consists of three layers.
The most superficial layer is called a mucous membrane, it is covered by a stratified squamous epithelium without glands; it is the peculiarly changed skin. This shell of vagina is of pink colouring, which, however, changes in some common conditions: during menstruation, pregnancy, inflammatory processes, etc. The cells of vaginal epithelium usually have cyclic changes in accordance with a menstrual cycle. There are four layers of cells of the vaginal mucous membrane distinguished by their functional and diagnostic significance: cells of superficial, or functional, layer (the area of cornification), cells of intermediate layer, parabasal cells, and typical basal cells (a deep layer). There are typical changes of all these cells during menstrual cycle. These changes are characteristic; they allow to judge about phases of menstrual cycle and about some hormonal disturbances, as well as about changes at pregnancy.
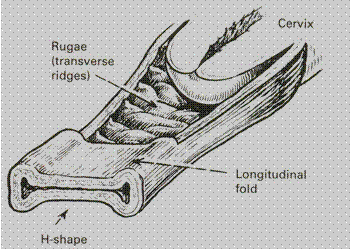
Fig. 9. The diagram of vagina
The middle layer of vaginal wall is muscular; it is highly developed in reproductive age. The musculature of vagina consists of three layers: internal — longitudinal, middle — circular, and outward — longitudinal. An outward layer is the most powerful.
The internal layer of vagina consists of a connective tissue membrane, directly transforming into the connective tissue, which separates the vagina from the neighbouring organs.
A vaginal tube from each side is surrounded by a paravaginal fat, which is mostly developed in the lateral parts.
Anteriorly: the vagina is related with the back of the bladder in its upper part, and in its lower part — with the urethra and fatty tissue behind and below the pubic symphysis.
Relations
Posteriorly: the vagina is covered with peritoneum in its most upper part, which separates the vagina from the loops of intestines. Below the peritoneum, it is related with the lowest part of the rectum and still lowers the perineal body, which separates the vagina from the anal canal. On each side the region of lateral fornix gives attachment to broad ligament and is related with the ureter and the uterine vessels above the ureter. (Fig. 10).
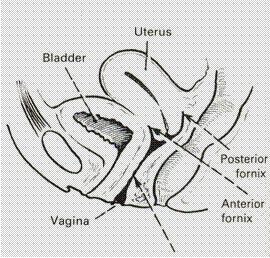
Fig. 10. Relations of the vagina with adjacent organs.
Uterus. The uterus is a hollow piriform muscular organ, which is situated in the pelvis between the bladder in front and the rectum behind. Its normal position is one of the anteversion and anteflexion. The uterus usually inclines to the right (dextrorotation) so that the cervix is directed to the left (levorotation) and comes in close relation with the left ureter. There is an obtuse angle between the corpus and the cervix of the uterus.
The uterus measures about 8 cm long, 5 cm wide at the fundus (from one tubal angle to another) and its walls are about 1. 25 cm thick. Its weight varies from 50 to 80 g. It has the following parts: the fundus, the body or the corpus, isthmus, cervix. (Fig. 11).
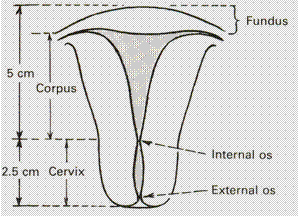
Fig. 11. The diagram of the uterus
Body, or corpus, of the uterus. The body proper is triangular and lies between the openings of the tubes and the isthmus. The part of the uterus that lies between the openings of the tubes is called the fundus of the uterus. The superolateral angles of the body of the uterus project outwards from the junction of the fundus and body and are called the cornua of the uterus. The uterine tube, round ligaments and ligaments of the ovary are attached to it. The cavity of the uterus is triangular on coronal section with the base above and the apex below. It measures about 3. 5 cm. There is no cavity in the fundus. The uterine wall consists of 3 layers from outside inwards:
• The 1st layer is perimetrium. It is a serous coat which covers the entire organ except for its lateral borders. The perimetrium is porously adhered to the underlying muscles.
• The 2nd layer is myometrium. It consists of thick bundles of smooth muscle fibres held by the connective tissues and arranged in 3 distinct directions: outer longitudinal, middle interlacing (consisting of circular and oblique fibres) and inner longitudinal.
• The 3d layer is endometrium. It is the mucous lining of the cavity. As there is no submucous layer, the endometrium is directly apposed to the muscle coat. Endometrium is lined by a cylindrical ciliary epithelium; epithelial cilia vibrate downward towards the external os of the cervical canal. The thickness of endometrium is 1-2 mm. The endometrium is composed of two layers separated from each other without distinct demarcation. They are a superficial layer (surface epithelium) and lamina propria. The superficial layer undergoes rhythmical changes according to a menstrual cycle, it is called a functional layer of endometrium. During the menstrual flow this layer of endometrial cells exfoliates and flows out together with menstrual blood. The functional layer of endometrium changes to the decidua during pregnancy.
The isthmus is a constricted part measuring about 0. 5 cm situated between the corpus (body) and the cervix of the uterus. It is limited above by the anatomical internal os and below by the histological internal os (Ashoff). The mucosal coat of the isthmus is identical to that of the endometrium; the structure of the wall of the isthmus is the same as the uterine wall.
Cervix is cylindrical in shape and measures 2. 5 — 3 cm. It extends from the isthmus and ends at the external os, which opens into the vagina after perforating its anterior wall. The portion of the cervix which lies in the vagina is known as vaginal part of the cervix, the part lying above the vagina is called supravaginal. The cervical canal is fusiform and measures about 2. 5 cm. Thus, the normal length of the uterine cavity including the cervical canal is about 6. 5 — 7 cm. Histologically it has inner lining of mucosa which is covered by columnar or high cuboidal epithelium and is made up of folds. The mucosa lies on the submucosa. The muscular coat is mostly a fibrous tissue and is arranged in a circular manner. Its blood supply comes from the cervical artery and vein which are the major branches of the uterine circulation.
Relations
Anteriorly: above the internal os, the body forms the posterior wall of the uterovesical pouch. Below the internal os, it is separated from the base of the bladder by a loose areolar tissue.
Posteriorly: it is covered with the peritoneum and forms the anterior wall of Douglas’ pouch containing coils of the intestine.
Laterally: the double folds of peritoneum form the broad ligaments of uterus, between which the uterine artery ascends up. About 1. 5 cm away at the level of internal os, a little closer to the left side there is a crossing of the uterine artery and the ureter. The uterine artery crosses the ureter from above and in front, just before the ureter enters the ureteric tunnel.
Fallopian tubes. These are a pair of long, thin, delicate tubes, length of each being approximately 10 cm, and situated in the medial three-fourths of the upper free margin of the broad ligament. (Fig. 12). They start from the cornu of the uterus and open into the peritoneal cavity just below and medially to the ovary on the same side. Each tube has two openings, one communicating with the lateral angle of the uterine cavity called uterine opening and measures 1 mm in diameter, the other is on the lateral end of the tube, called pelvic opening, and measures about 2 mm in diameter.

Fig. 12. Fallopian tube, ovary and suspending ligaments: A. –Ampula of the fallopian tube; M- Broad ligament; E-Epioophoron, I- Isthmus of the tube; Ov-ovary; L-Proper ligament of the ovary; F- fimbriae of the tube.
Four parts are distinguished in each tube proceeding from uterus to ovary:
• intramural or interstitial lying in the uterine wall and measures 1. 25 cm in length and 1 mm in diameter;
• isthmus is almost straight and measures about 2. 5 cm in length and 2. 5 mm in diameter;
• ampulla — a tortuous part measuring about 5 cm in length which terminates widely;
• infundibulum measuring about 1. 25 cm long with a maximum diameter of 6 mm.
The abdominal ostium is surrounded by a number of radiating fimbriae, one of which is longer than the rest and is attached to the outer pole of the ovary called the ovarian fimbria.
Every tube consists of 3 layers: serous, muscular and mucous.
The ovaries. The ovaries are double sex glands or gonads in a female which are responsible for germ cell maturation, storage and its release, and steroidogenesis. Each ovary is oval in shape and pinkish grey in colour and the surface is scarred during the reproductive period. It measures about 3 cm in length, 2 cm in width and 1 cm in thickness.
The ovaries are extraperitoneal structures. The ovary is inserted by its one end into the posterior leaf of the broad ligament while the rest of it remains uncovered by the peritoneum and is suspended in the pelvic cavity. The ovary is attached to the posterior layer of the broad ligament by the mesovarium, to the lateral pelvic wall — by the infundibulopelvic ligament and to the uterus — by the ovarian ligament.
Two surfaces are distinguished in the ovary: internal, turned toward the abdominal cavity (facies medialis), and external, turned to the wall of pelvis (facies lateralis). There are two ends — uterine and tubal, and two edges — protuberant (free) and mesenterial, to which mesentery of the ovary is attached. The vessels and nerves enter the ovary at the point where the latter is inserted into the posterior leaf of the broad ligament (hylus of the ovary).
The free surface of the ovary is covered by a monostratal cubical epithelium which makes the first, superior layer of the ovary. Under the epithelium, the proteinous layer (tunica albuginea) is disposed; it consists of connective tissue which fibres are parallel to the ovarian surface. The thickness of the proteinous layer does not exceed 0. 1 mm. Under the proteinous layer of the ovary its basic layer lies — the cortex of the ovary, under which the fourth layer, named medulla, is situated. The cortex of the ovary consists of follicles in different stages of development. The medulla of the ovary consists of soft connective tissue, which contains a lot of blood and lymphatic vessels and nerves.
Ligamentous apparatus of the uterus and ovaries
The normal position of the uterus, tubes, and ovaries in the pelvic cavity mainly depends on the ligaments and the muscles of the pelvic floor. These form the suspensory, immobilizing, and supporting structures.
The suspending structure of the internal genitalia comprises the following (Fig. 13):
• Round ligaments (lig. rotundum, s. lig. teres). Round ligaments extend from the angles of the uterus (somewhat anteriorly and below the insertion of the fallopian tubes). Each runs in the anterior leaf of the broad ligaments to the inguinal canal, through which it passes to terminate in the connective tissue of the mons pubis and the labia majora. The round ligaments pull the uterus bottom anteriorly (anteflexion). During pregnancy the round ligaments thinned and elongate. There is a small branch of inferior epigastric artery (or a. spermatica externa) inside each round ligament, called artery of a round ligament which supplies round ligament and labia majora.
• Broad ligaments (lig. latum). These are twin-leaves of the peritoneum running from the uterus sides to the lateral walls of the pelvis. The superior portion of the broad ligament contain the fallopian tubes, while the posterior leaves suspend the ovary. The vessels and nerves pass in the connective tissue between the leaves.
• Sacrouterine ligaments (lig. sacro-uterinum). These ligaments originate from the posterior surface of the uterus at the point where the isthmus transforms into the cervix, and pass backward to embrace the rectum on both sides and are attached to the anterior surface of the sacrum. These ligaments tend to keep the cervix posteriorly. During labor the round and sacrouterine ligaments oppose the displacement of the uterus.
• Proper ligaments of the ovaries (lig. ovarii proprium). These originate at the fundus of the uterus, posteriorly and below the insertion of the fallopian tubes, and extend to the ovaries.

Fig. 13. Ligament structure of internal genital organs: 1-Round ligament; 2- Proper ligaments of the ovaries (lig. ovarii proprium); 3- Mesosalpinx; 4-oviduct; 5-epioophoron; 6- ductus epioophori longitudinalis; 7-ampula of the tube; 8-tubal fimbriae; 9- infundibulum; 10-ovary; 11- broad ligament; 12-plica recto-uterina; 13- vagina; 14- peritoneum; 15-cervix of the uterus; 16- corpus of the uterus.
The immobilizing system of the uterus is composed of connective-tissue bands incorporating smooth muscles. They extend from the inferior portion of the uterus anteriorly to the bladder and further to the symphysis, to the lateral walls of the pelvis (cardinal ligaments), and posteriorly to form a connective-tissue framework of the sacrouterine ligaments.
The supporting system comprises the muscles and the fasciae of the pelvic floor.
Blood supply, lymphatic and nervous system of the female reproductive organs
Blood supply
The circulatory system of pelvic organs is highly developed. It particularly refers to the vascular system of the uterus, which is in great need of rich blood supply during pregnancy.
Female genital organs get blood from two sources — from internal iliac artery and from internal ovarian artery.
Blood Supply of the external genital organs
Arteries supplying the external genitalia (Fig. 14) are:
• Branches of internal pudental artery — the chief being labial, transverse perineal, artery of the vestibular bulb and deep and dorsal arteries to the clitoris.
• Branches of femoral artery — superficial and deep branches of external pudendal artery.

Fig. 14. Arteries of female pelvic organs.
1 -abdominal aorta; 2-inferior mesenteric artery; 3-common iliac artery; 4-external iliac artery; 5- internal iliac artery; 6- superior gluteal artery; 7- inferior gluteal artery; 8- uterine artery; 9- umbilical artery; 10- vesical arteries; 11- vaginal artery; 12- internal pudendal artery; 13- perineal artery; 14- inferior rectal artery; 15- artery of the clitoris; 16- middle rectal artery; 17- uterine artery; 18- tubal branch; 19- ovarian branch; 20- ovarian artery; 21- median sacral artery.
Blood Supply of the Vagina
Blood supply of the vagina is carried out due to the branches of internal iliac artery (a. iliaca interna, s. hypogastrica) and vaginal artery (r. servicovaginalis a. uterinae), the latter being a descending branch of uterine artery. The vaginal artery branches off the uterine artery at the level of the uterine cervix and proceeds along the lateral walls of the vagina supplying mainly its superior part. The middle part of the vagina is mainly provided with the inferior vesical artery (a. vesicalis inferior), middle rectal artery (a. haemorroidalis media, s. a. rectalis media), and internal pudendal artery (a. pudenda interna). There are some small branches of internal pudendal artery passing to the entrance of vagina (aa. labiales posteriores).
Blood Supply of the Uterus
It is provided by two uterine arteries, one on each side. The artery arises directly from the anterior part of the internal iliac artery or from the superior vesical artery. Another source of blood supply is ovarian and vaginal arteries which the uterine arteries anastomose with.
Every uterine artery passes to the uterus through the perimetric fat between two leaves of broad ligament, and, passing in front of the ureter, approaches the internal uterine os, where it is divided into two branches — ascending and descending. The right uterine artery anastomoses with the left one in the middle of the uterus. An ascending branch goes up as a cork-screw joining near the tubal corner of the uterus with the branch of the external spermatic artery, which supplies the round ligament.
Moreover, it branches off to the fundus of the uterus, to the fallopian tube, and to the ovary.
A descending branch of the uterine artery supplies the cervix of the uterus and upper part of the vagina; it anastomoses with the same branches from the opposite side.
Blood Supply of the Fallopian Tubes
The arterial blood supply is provided by the uterine and ovarian arteries. Venous drainage is through the pampiniform plexus into the ovarian veins.
Blood Supply of the Ovary
The arteries supplying the ovary are the ovarian artery, a branch of the abdominal aorta, which anastomoses with the ovarian branch of the uterine artery.
Veins of Genital Organs
The vein system of pelvic organs is highly developed (Fig. 15). The common extension of its vessels considerably exceeds the extension of the arterial system. Blood from the uterus outflows through veins forming uterine plexus (plexus uterinus) in three directions:
• Ovarian vein (from the ovary, tube and upper segment of the uterus);
• Uterine vein (from lower segment of the uterus and upper part of the cervix);
• Iliac vein (from the lower part of the cervix and from vagina).
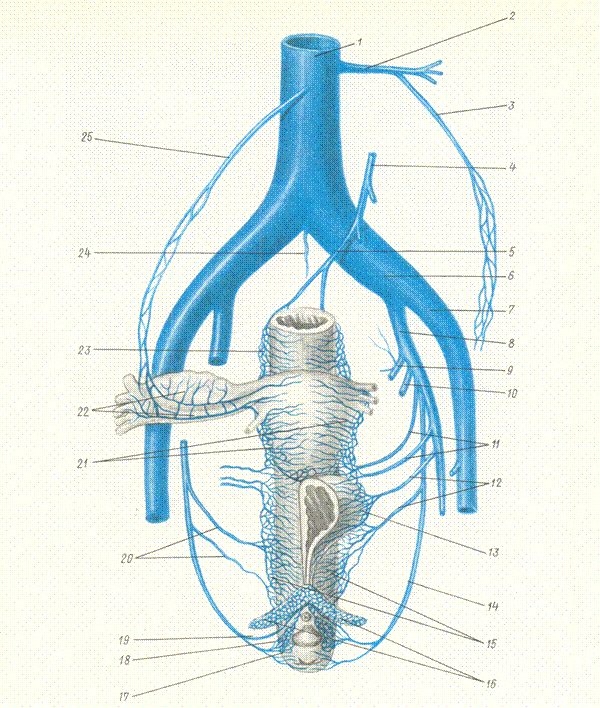
Fig. 15. Veins of female pelvic organs. 1- vena cava inferior; 2-left renal vein; 3- left ovarian vein; 4- inferior mesenteric vein; 5- superior rectal vein; 6- common iliac vein; 7- external iliac vein; 8- internal iliac vein; 9- superior gluteal vein; 10- inferior gluteal vein; 11- uterine veins; 12- vesical veins; 13-vesical veniplex; `14- internal pudendal vein; 15- vaginal veniplex; 16- crura clitoridis; 17- inferior rectal vein; 18 – bulbus vestibule; 19- vein of the clitoris; 20- vaginal veins; 21- uterine veniplex; 22 – pampiniform veniplex; 23 – rectal veniplex; 24 – median sacral vein; 25 – right ovarian vein.
There are anastomoses between uterine plexus, veins of urinary bladder and plexus rectalis.
Blood from the ovary is drained through pair of ovarian veins which correspond to arteries. They start from pampiniform plexus, pass through suspensory ligament of the ovary, empty into the vena cava inferior (right) and into the left renal vein (left).
Vaginal veins form plexuses on the lateral walls of the vagina which anastomose with venous system of external genitalia and adjacent organs of the pelvis. The outflow of blood from these plexuses takes place into internal iliac vein.
Veins of external genital organs
Veins form plexuses and drain into the internal pudendal vein, vesicular or vaginal venous plexus, and long saphenous vein. Varicosities in pregnant are not very rare and may rupture spontaneously during pregnancy forming hematoma or visible hemorrhage. On the other hand, varicosities may rupture during childbirth causing profuse hemorrhage.
Lymphatic system
The lymphatic system (Fig. 16) of pelvic organs consists of lymphatic vessels interlacing in a form of a compound network and extending from external genital organs upwards along the retroperitoneal space up to the lumbar areas. The lymphatic system includes multiple lymphatic nodes collecting lymph which inflows to them through lymphatic vessels.
The lymphatic plexuses of mucous and serous layers of the uterus are distinquished. Plexuses in the muscular layer of the uterus form due to the perforated vessels which are a connecting link of vascular networks of mucous and serous layers of the uterus. The lymphatic vessels of the uterine cervix are connected with the lymphatic vessels of vagina, being continuation of vessel networks of mucous membrane of vagina.
The lymphatic nodes of pelvis are subdivided into parietal and visceral.

Fig. 16. 1 – n. l. lumbales; 2- n. l. iliaci communis; 3 – n. l. sacrales; 4 – n. l. rectalis; 5- n. l. iliaci externa; 6 – uterus; 7 – oviduct; 8 – n. l. iliaci initerni; 9 –a. uterina; 10 - a. iliaca interna; 11 – m. iliacus; 12 – a. ovarica; 13 – ureter; 14 – m. guadratus lumborum.
|
|
|
