 |


Физиология мышечного сокращения
|
|
|
|
Один из известных современных физиологов И.А. Аршавский писал, что «жизнью человека, его ростом и развитием правит мышечная активность». Мышечная система выполняет ряд важных функций. Динамическая функция состоит в движении крови по кровеносным сосудам (сердечная мышца), перемещении содержимого полых внутренних органов (гладкие мышцы их стенок), движении частей тела или всего тела в пространстве (скелетные мышцы). Работа скелетных мышц способствует активации и тренировке всех физиологических систем организма, особенно сердечно-сосудистой и дыхательной, повышая резервные возможности организма.
Статическая функция обеспечивает определенный тонус стенок внутренних органов (гладкие мышцы), поддержание позы, удерживание груза (скелетные мышцы).
Рецепторная функция связана с наличием в мышечной ткани многочисленных проприорецепторов, поставляющих центральной нервной системе информацию о свойствах внутренней среды и состоянии скелетной мускулатуры. Информация от рецепторов мышечной ткани способствует саморегуляции параметров внутренней среды организма, уровня активности, тонуса мышц, координации движений.
Метаболическая (обменная) функция заключается в том, что мышцы служат депо гликогена и белка, запасы которых расходуются при функциональном напряжении организма. Энергия АТФ, обеспечивающая мышечные сокращения, только на 40% расходуется на совершение механической работы, остальная ее часть рассеивается в виде тепла, поэтому мышцы являются важнейшим компонентом системы терморегуляции.
Мышечная ткань состоит из мышечных клеток – миоцитов. Особенностью миоцитов является наличие в цитоплазме связывающего кислород пигмента миоглобина и особых органоидов – миофибрилл, состоящих из молекул сократительных белков, основные из которых – актин и миозин. По строению различают три вида мышечной ткани – гладкую, сердечную (миокард) и поперечнополосатую. Гладкие мышцы составляют стенки внутренних органов, миокард выделяется в отдельную группу, поскольку имеет особое строение, сочетающее черты гладких и поперечнополосатых мышц, поперечнополосатые образуют скелетную мускулатуру (рис.13).
|
|
|
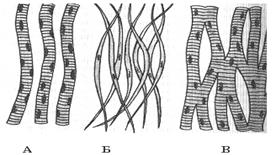
| Рис. 13. Разновидности мышечной ткани: А – поперечно-полосатая; Б – гладкая; В - миокард |
Мышечные клетки скелетной мускулатуры (мышечные волокна) – многоядерные клетки вытянутой формы, содержащие большое количество параллельно расположенных миофибрилл. Каждая миофибрилла разделена на сегменты (саркомеры) длиной около 2,5 мкм поперечными белковыми перегородками – Z-пластинками. Границы саркомеров миофибрилл одного волокна совпадают, это создает заметную по микроскопом поперечную исчерченность.
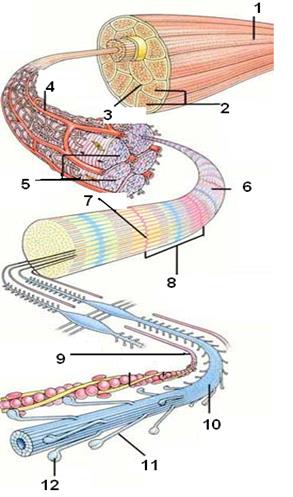
| Рис. 14. Строение мышцы: 1 – мышца; 2 – группы мышечных волокон; 3 - эндомизий; 4 – мышечное волокно с кровеносными сосудами; 5 – миофибриллы; 6 – отдельная миофибрилла; 7 - Z-пластинка; 8 – саркомер; 9– актиновая нить; 10 – миозиновая нить; 11 – актомиозиновый мостик; 12 – головка актомиозинового мостика |
Миофибриллы состоят из тонких нитей актина, прикрепленных к Z-пластинкам, и толстых миозиновых нитей, расположенных в центральной части саркомера между актиновыми нитями. Миозиновые нити имеют выступающие мостики, которые могут связываться с актиновыми нитями. Каждый мостик состоит из «шейки» и «головки», на которой расположены участки связывания АТФ (рис.14). Каждая миофибрилла окружена саркоплазматическим ретикулумом, в котором различаются поперечные трубочки, проходящие по границам саркомеров (Т-система) и сеточку, окружающую центральную часть саркомера (L – система). Внутри ретикулума находятся запасы ионов кальция, концентрация которого здесь в 10000 раз выше, чем в цитоплазме.
|
|
|
Различают красные, белые и промежуточные волокна. Красные волокна - тонкие, богатые саркоплазмой, гликогеном, миоглобином, митохондриями, содержат липидные включения, миофибрилл относительно немного. В них интенсивно идут окислительные процессы, они способны к длительному сокращению. Белые - толстые, богаты миофибриллами, митохондрий и миоглобина меньше. Они сокращаются быстрее, чем красные, но быстрее и утомляются. У человека мышцы содержат все типы волокон, те или иные преобладают в зависимости от функции мышцы.
В основе мышечного сокращения лежит взаимодействие между актином и миозином, при котором используется энергия, выделяющаяся при гидролизе АТФ. Катализатором реакции гидролиза АТФ является миозин. Механизм мышечного сокращения в настоящее время объясняется с позиций теории «скользящих нитей», разработанной американскими учеными Хаксли и Хансоном в 50-60 гг. 20-го века. Последовательность процессов, приводящих к мышечному сокращению, такова:
1. Нервный импульс по аксону мотонейрона достигает нервно-мышечного синапса, называемого концевой пластинкой. Ацетилхолин, выделяющийся из пресинаптического окончания, вызывает деполяризацию постсинаптической мембраны мышечного волокна и формирование потенциала концевой пластинки (ВПСП).
2. Под действием потенциала концевой пластинки на прилежащих участках мембраны миоцита возникает ПД, который распространяется по Т- системе на L – систему и вызывает выход ионов кальция в цитоплазму.
3. Кальций взаимодействует с тонкими нитями таким образом, что на молекулах актина освобождаются участки, которые могут связываться с миозиновыми мостиками.
4. «Головка» миозиновой нити присоединяется к активному участку актиновой нити и одновременно связывается с молекулой АТФ и катализирует ее гидролиз, в результате чего выделяется энергия. Энергия затрачивается на образование акто-миозиновой связи и изменение пространственной структуры мостика, приводящее к его укророчению.
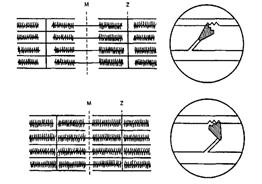
5. В результате укорочения мостика актиновая нить передвигается к центру саркомера - это напоминает гребок весел. Каждый такой «гребок» укорачивает саркомер на 1 %, за 0,1 с таких движений может быть до 50 (рис.15).
|
|
|
6. Когда к данному участку подходит следующая молекула АТФ, актомиозиновый мостик разрывается и замыкается вновь на следующем участке актиновой нити. Если АТФ недостаточно, формируется стойкое сокращение – контрактура. Исчезновением энергетических запасов можно объяснить развитие «трупного окоченения», которое проходит в результате посмертного разрушения миофибрилл.
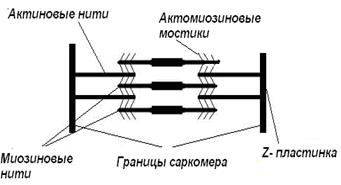
| 
|
| А | Б |
Рис.15. Схема строения саркомера (А) и замыкания актомиозиновых мостиков (Б)
Если одновременно сокращаются все участки мышцы, то ее длина укорачивается (изотоническое сокращение). Сочетание укорочения одних участков мышечного волокна с растяжением других приводит к изометрическому сокращению (длина мышцы не меняется, а тонус увеличивается).
Силу мышечного сокращения можно измерить величиной груза, который мышца может поднять при сокращении. Она вычисляется делением массы максимального груза, который может поднять мышца (кг), на площадь ее поперечного сечения (см2). Так, абсолютная сила икроножной мышцы человека 5,9 кг/см2, двуглавой мышцы плеча 11,4 кг/см2, трехглавой мышцы плеча 16,8 кг/см2. Для одного мышечного волокна сила составляет 0,1-0,2 г.
Во время физических нагрузок увеличивается приток крови к мышцам, усиливаются энергетические процессы, что создает условия для образования новых миофибрилл. Регулярные упражнения приводят к утолщению (гипертрофии) мышечных волокон. При очень сильных нагрузках в мышцах могут возникать микроразрывы, что стимулирует образование новых мышечных волокон за счет стволовых клеток, присутствующих в мышечной ткани (гирперплазия мышц).
Важным элементом мышечной ткани являются нервно-мышечные веретена, выполняющие рецепторную функцию (проприорецепторы мышц).
| Такие веретена образованы видоизмененными миниатюрными мышечными волокнами (длина 2мм, диаметр около 0,1 мм), расположенными между настоящими волокнами, заключенными в капсулу из соединительной ткани и оплетенными чувствительными и двигательными нервными окончаниями. | 
|
| Рис.16. Мышечное веретено |
Чувствительные окончания возбуждаются при растяжении веретена, двигательные являются отростками гамма – мотонейронов. Подобные рецепторные образования имеются и в сухожилиях – сухожильные органы Гольджи. Нервно-мышечные веретена и сухожильные комплексы обеспечивают вторичную афферентацию и информируют ЦНС о состоянии двигательного аппарата и результатах рефлекторных двигательных реакций.
|
|
|
Лекция 9. СПИНАЛЬНЫЙ И СТВОЛОВОЙ УРОВНИ РЕГУЛЯЦИИ ДВИЖЕНИЙ
Спинномозговой уровень регуляции движений
Основные функции спинного мозга – рефлекторная и проводниковая. В передних рогах спинного мозга расположены мотонейроны, образующие сегментарные двигательные центры.
Рефлексы, центры которых расположены на уровне спинного мозга, называются собственными рефлексами спинного мозга, или спинальными рефлексами. Эти рефлексы теоретически могут осуществляться и без участия головного мозга.
1. Миотатические рефлексы, или рефлексы на растяжение. Рецепторы - проприорецепторы мышц и сухожилий, адекватным стимулом для которых является расслабление или растяжение мышцы. Дуги таких рефлексов имеют минимальное число синаптических переключений. Возбуждение с чувствительного нейрона может сразу передаваться на эфффекторный мотонейрон спинного мозга. Центры располагаются на уровне 3-х соседних сегментов. Возбуждение центров, обеспечивающих сокращение мышц – сгибателей вызывает одновременную индукцию торможения в нейронах, иннервирующих мыщцы-антагонисы (разгибатели) посредством латерального торможения. Это явление называется реципрокной иннервацией мышц-антагонистов. Исследование сухожильных рефлексов имеет диагностическое значение для определения состояния сегментарного аппарата спинного мозга: коленный рефлекс замыкается на уровне 2-4 поясничных сегментов, рефлекс с ахиллова сухожилия – на уровне 1-2 крестцовых, локтевые рефлексы – на уровне 5-6 шейных сегментов.
Тонус мышц, регулируемый спинным мозгом, развивается в основном в мышцах- сгибателях. Спинальный тонус формируется еще во внутриутробном периоде. Он необходим плоду для принятия эмбриональной позы, позволяющей ему занимать как можно меньше места в организме матери. Новорожденному ребенку спинальный сгибательный тонус нужен для уменьшения потери тепла.
2. Сгибательные рефлексы – защитные. Возникают при раздражении болевых и механорецепторов поверхности тела и проявляются сокращением мышц-сгибателей. Такие рефлексы можно наблюдать у спинальной лягушки. У новорожденного и ребенка до 3-х месячного возраста можно наблюдать хватательный рефлекс, рефлекс Моро – симметричные обхватывающие движения ручек. У взрослого человека такие рефлексы включены в двигательные реакции при боли.
|
|
|
3. Перекрестные разгибательные рефлексы, возникающие одновременно со сгибательными и выражающиеся в разгибании конечности противоположной стороны. Физиологическим смыслом таких рефлексов является обеспечение опоры.
4. Спинальные двигательные автоматизмы. На уровне спинного мозга имеются нейронные цепи, являющиеся субстратом врожденных двигательных программ – ходьбы, плавания, чесания. Так, у собаки с перерезанным спинным мозгом можно вызвать ритмические чесательные движения лапы раздражением кожи. У ребенка сразу после рождения наблюдается шаговый автоматизм, при погружении в воду он совершает ритмические «плавательные» движения.
Функции спинного мозга контролируются вышележащими отделами ЦНС. Чем выше уровень развития организма, тем более выражен этот контроль. При нарушении связей спинного и головного мозга наблюдается временное исчезновение спинальных рефлексов – спинальный шок. У лягушки он продолжается несколько минут, у собаки – несколько часов, у обезьяны - несколько недель, у человека – несколько месяцев. Симптомы, наблюдающиеся при полном поперечном разрыве спинного мозга, иллюстрируют его функции:
- полный необратимый паралич произвольных движений мышц, иннервируемых сегментами, расположенными ниже повреждения;
- полная необратимая потеря осознаваемой чувствительности в этих областях тела;
- полная обратимая утрата спинальных рефлексов, центры которых расположены в сегментах ниже повреждения.
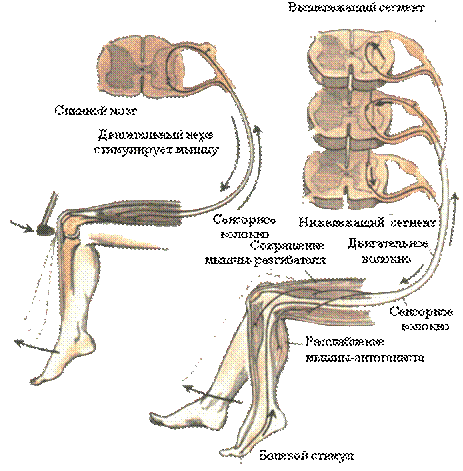
| Рис.17. Сухожильный и сгибательный спинальные рефлексы |
Стволовой уровень регуляции движений
Стволовые соматические рефлексы можно разделить на несколько групп.
Рефлексы пищевого поведения и защитные рефлексы – глотание, сосание, жевание, рвотный рефлекс, чихание, кашель, мигательный рефлекс. Запускаются при раздражении рецепторов слизистой оболочки ротовой и носовой полости, иннервируемых чувствительными волокнами блуждающего, языкоглоточного и тройничного нервов. Центрами этих рефлексов являются ядра тройничного, языкоглоточного, лицевого, блуждающего, добавочного и подъязычного нервов в продолговатом мозге и мосту. Эти рефлексы могут осуществляться без участия вышележащих отделов ЦНС и наблюдаются даже у анэнцефалов.
Тонические рефлексы – проявляются в тоническом сокращении мышц-разгибателей. Возникают при раздражении рецепторов вестибулярного аппарата, проприорецепторов мышц шеи и туловища. Надсегментарными центрами таких рефлексов является вестибулярные ядра Дейтерса и Швальбе, аксоны которых формируют вестибулоспинальные пути, заканчивающиеся на интернейронах и мотонейронах спинного мозга. Эти ядра оказывают возбуждающее действие на мотонейроны разгибателей и тормозное – на мотонейроны сгибателей. Центром, тормозящим активность этих ядер, является красное ядро. При перерезке ствола на уровне моста и разобщении красного и вестибулярного ядер наблюдается децеребрационная ригидность, проявляющаяся в развитии универсального разгибательного тонуса: у животного (кошки) вытягиваются передние и задние конечности, поднимается хвост, разгибается шея и голова запрокидывается назад. Согнуть конечности в суставах практически не удается (рис.18).

| Рис. 18. Децеребрационная ригидность при разобщении красных и вестибулярных ядер |
В онтогенезе разгибательный тонус появляется позже сгибательного, так как структуры продолговатого мозга созревают позже спинного мозга. Только на втором месяце жизни у детей появляется рефлекс Ландау - ребенок, лежа на животе, поднимает голову, опираясь на предплечья, а позже на кисти рук. На 5-6 месяце ребенок разгибает ноги и приподнимает таз, что создает условия для ползания. При этом увеличивается пространство, видимое глазом, обогащается жизненный опыт ребенка.
Магнусовские рефлексы - названы по имени голландского ученого Р.Магнуса, описавшего эти рефлексы в 20-х годах 20 века. Эти рефлексы разделяются на 2 группы – статические и статокинетические. Центры – вестибулярные, красные ядра и некоторые другие стволовые структуры.
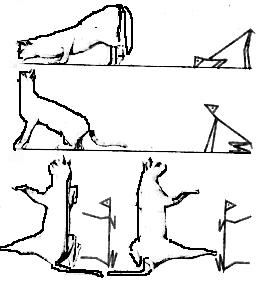
Статические рефлексы – это рефлексы позы и рефлексы положения. Рефлексы позы возникают при поворотах и наклонах головы (раздражение вестибулярных рецепторов и проприорецепторов шеи) и выражаются в изменении тонуса мышц - сгибаталей и разгибателей, что в целом обеспечивает равновесие и удобную позу. Так, при наклоне головы у животного передние конечности сгибаются, а задние – разгибаются (удобнее поднимать пищу с земли); при запрокидывании головы задние конечности сгибаются, передние разгибаются (удобно тянуться за пищей, находящейся над головой). У человека такие рефлексы наблюдаются в первое полугодие жизни, а затем затормаживаются развивающимися вышележащими центрами: у младенца при повороте головы сгибаются конечности на стороне, противоположной повороту, и разгибаются с той стороны, куда поворачивается голова. У взрослого человека такие рефлексы включаются в сложные двигательные программы (например, при спортивных движениях – группировка при прыжках в воду, кувырках и т.д.).
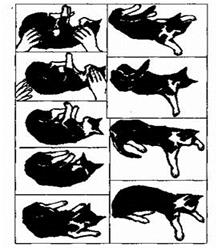
Рефлексы положения возникают при изменении положения головы в пространстве. «Нормальным» для большинства животных является положение «теменем кверху». При отклонениях от этого положения происходит рефлекторное восстановление нормальной ориентации (если положить на спину лягушку, кролика или морскую свинку, то они тот час же перевернутся обратно). У более высокоразвитых животных и человека центры этих рефлексов тормозятся вышележащими уровнями, поэтому они могут длительное время сохранять разные положения тела, но кошка, падая с высоты, приземляется на лапы (рис.19).
| 
|
| А | Б |
Рис.19. Магнусовские рефлексы
А – рефлексы позы; Б – рефлексы положения
Статокинетические рефлексы развиваются при ускоренных прямолинейных и вращательных движениях. Мы можем наблюдать их у себя при катании на каруселях и качелях, при поездках на транспорте, подъеме и спуске в лифте. Возникают при раздражении вестибулярных и зрительных рецепторов. В начале вращения туловище отклоняется в сторону, противоположную вращению, голова и глазные яблоки медленно отклоняются в сторону, противоположную вращению, затем быстро возвращаются в сторону вращения (нистагм). При замедлении и остановке движения туловище медленно отклоняется в сторону вращения, направление нистагма меняется. Подобные рефлексы имеют вегетативный компонент в виде возбуждения симпатических (тахикардия, повышение потоотделения) или парасимпатических центров (побледнение, тошнота, рвота). Регулярные воздействия на вестибулярный аппарат снижают возбудимость вестибулярных рецепторов (адаптация), и рефлексы становятся менее выраженными. При движении вверх и вниз наблюдается «лифтный» рефлекс – при подъеме повышается тонус сгибателей конечностей, при спуске – тонус разгибателей. Биологическая роль таких рефлексов заключается в поддержании равновесия тела и облегчении зрительной ориентации во время движения.
Ориентировочные рефлексы - возникают при действии новых зрительных и слуховых раздражителей и проявляются в реакциях настораживания – остановка текущей деятельности, повышение тонуса скелетной мускулатуры, поворот глаз, головы и туловища в сторону раздражителя. Эти реакции включают вегетативный компонент преимущественно симпатической направленности – расширение зрачков, учащение сердцебиений, повышение АД. Эти реакции способствуют энергетическому обеспечению защитного поведения в случае, если незнакомый раздражитель окажется угрожающим. Основными центрами ориентировочных реакций являются ядра четверохолмия, поэтому и рефлексы часто называют четверохолмными.
Кроме осуществления собственной рефлекторной деятельности, спинальные и стволовые центры принимают участие в реализации движений, формирующихся при участии ДЦ вышележащих уровней ЦНС.
Лекция 10. ПИРАМИДНАЯ И ЭКСТРАПИРАМИДНАЯ СИСТЕМЫ ДВИГАТЕЛЬНОГО КОНТРОЛЯ
Нисходящие регулирующие влияния структур конечного мозга на сегментарные двигательные центры осуществляется через две системы –пирамидную и экстрапирамидную. Обе этих системы берут начало в моторной коре, занимающей прецентральную извилину и задние отделы верхней и средней лобных извилин.
Пирамидная система представлена гигантскими пирамидными нейронами Беца 5-го слоя прецентральной извилины и пирамидными нисходящими путями, образованными аксонами этих клеток. Пирамидные пути заканчиваются непосредственно на мотонейронах сегментарных ДЦ. Большинство пирамидных волокон переходит на противоположную сторону (боковые пирамидные пути – на уровне продолговатого мозга, передние – на уровне спинного мозга) (рис.20).
Эта система обеспечивает точный и быстрый контроль произвольных движений. Кора прецентральной извилины организована соматотопически: за сокращения разных групп мышц «отвечают» строго определенные участки коры. Еще в 1870 году, во время прусско-датской войны, немецкий военный хирург Т.Фрич заметил, что при обработке проникающих ран определенных участков головы наблюдаются подергивания мышц конечностей. Он сообщил об этом факте физиологу Э. Гитцигу, тот поставил серию опытов, в результате чего была создана карта представительства мышц в моторной коре – «двигательный гомункулюс». Наибольшую площадь занимают участки коры в дорсолатеральной части прецентральной извилины, связанные с движениями пальцев рук, кистей, языка, мышц лица, и значительно меньшую – представительства туловища и нижних конечностей в медиальной части прецентральной извилины. Неравномерность распределения двигательной коры между различными частями тела обусловлена тем, что мышцы, обеспечивающие тонкие и точные движения, связанные с трудовой деятельностью и речью, иннервируются большим количеством мотонейронов (двигательные единицы мелкие). Этими мотонейронами «управляют» соответственно многочисленные корковые двигательные клетки.
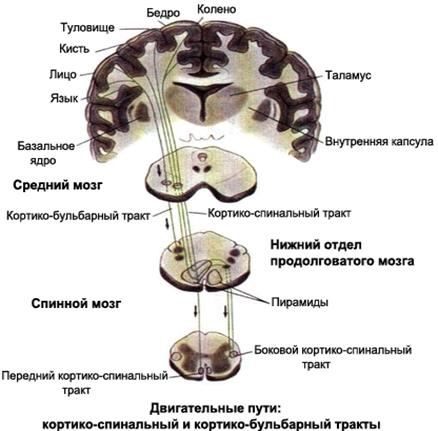
| Рис. 20. Пирамидная система регуляции движений |
В настоящее время установлено, что и двигательные, и сенсорные зоны коры имеют колончатое строение. Корковая колонка представляет собой вертикально ориентированный ансамбль примерно из 100 нейронов, проходящий через все слои коры. Каждая такая колонка связана с двигательными единицами определенных движений, например, сгибания руки в лучезапястном суставе. Между моторными зонами обоих полушарий имеются двусторонние связи. Так, при одностороннем поражении моторной коры происходит постепенная компенсация утраченных движений, при двусторонних поражениях произвольные движения не восстанавливаются. Пирамидная система регуляции включается под влиянием интегративно-пусковых зон лобной коры, в которых формируется стратегия и план движения. При электроэнцефалографии в лобных зонах еще до начала движения регистрируется повышение электрической активности - «потенциалы ожидания и готовности».
Экстрапирамидная система осуществляет регуляцию общего уровня двигательной активности, тонуса мускулатуры, контролирует автоматизированные движения (мимику, бег, ходьбу, плавание и т.д.). Экстрапирамидная система включает базальные ядра, черную субстанцию и красное ядро среднего мозга, мозжечок, корковые структуры. Особенность этой системы является большое количество переключений нисходящих путей на разных уровнях ЦНС, прежде чем сигнал достигнет сегментарных двигательных центров.
Важнейшим компонентом экстрапирамидной системы является стриопаллидарная система – хвостатое ядро, скорлупа, бледный шар. Хвостатое ядро и скорлупа - это неостриатум, бледный шар – палеостриатум. Полосатое тело получает афферентацию от коры мозга (двигательные зоны коры, соматосенсорная кора и др. области как своего, так и противоположного полушария), таламуса, черной субстанции среднего мозга, других ядер ретикулярной формации. Неостриатум посылает эфференты к бледному шару, через который связывается практически со всеми отделами ЦНС. Значение стриарной системы в целом отчетливо выявляется в опытах со стриарными и таламическими животными. У стриарных животных (кошек) разрушена КБП, но сохранена стриопаллидарная система. Такие кошки способны умываться, находить и поедать пищу, но двигательная активность у них снижена. Таламические кошки уже не умываются, питаются искусственно, двигательная активность у них повышена. Они могут выполнять простые рефлексы, если их вызывать изолированно: облизывают пищу, если поднести ее ко рту, но не захватывают, если пища во рту – жуют, но не глотают, а проглатывают пищу, если поместить ее на корень языка. Видимо, одна из функций полосатых тел – обеспечение необходимой последовательности движений при сложных безусловных рефлексах. Стриарные приматы гораздо менее «способные», они малоподвижны, могут только сидеть, живут без коры только около 2 недель.
Функции отдельных элементов стриопаллидарной системы изучаются с помощью методов разрушения и раздражения в экспериментах на животных. Мы рассмотрим двигательные функции, хотя стриопаллидарная система участвует также в регуляции эмоциональных состояний, процессах научения и памяти. Повреждение полосатого тела вызывало у животных безудержное стремление двигаться вперед, резко увеличивалась общая двигательная активность, но последовательность движений и координация были нарушены, появлялись насильственные мимические движения, тремор, судороги. У человека с повреждением полосатых тел наблюдаются гиперкинезы типа торсионного спазма и хореи. Торсионный спазм – вращательное штопорообразное движение туловища. Хорея – быстрые изменения движений конечностей, туловища, лица, языка и мягкого неба. Раздражение хвостатых ядер, напротив, приводило к остановке начавшихся произвольных движений. Тормозящие влияния хвостатого ядра являются условием перехода одного действия в другое.
Повреждение бледного шара, напротив, приводит к снижению двигательной активности, гипомимии, тремору головы и конечностей, который исчезает в покое и усиливается при движениях. Те же симптомы наблюдаются при недостаточности функции черной субстанции среднего мозга. Подобный комплекс симптомов называется паркинсонизмом.
Таким образом, можно заключить, что бледный шар инициирует движения, делает их сильными и размашистыми. Его активность регулируется тормозными влияниями полосатого тела. В свою очередь, полосатое тело испытывает тормозящие влияния со стороны черной субстанции среднего мозга.
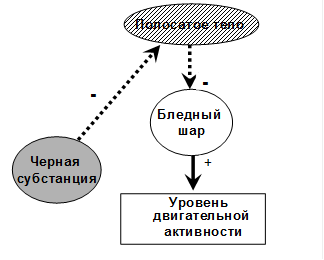
| Рис.21. Взаимодействие компонентов стриопаллидарной системы |
Другим важным элементом экстрапирамидной системы является мозжечок. Он, как и стриопаллидарная система, является хранилищем центральных моторных программ. Мозжечок связан со всеми отделами ЦНС. В ответ на корковую команду он выбирает нужную программу и «отправляет» ее в КБП, одновременно подготавливая сегментарные моторные центры к ее выполнению. На основании информации, получаемой от проприорецепторов, вестибулярных и других сенсорных систем, он регулирует тоническую активность мотонейронов, уровень их возбудимости, обеспечивает быстрый переход от сгибания к разгибанию, и наоборот. В целом можно сказать, что он обеспечивает синергию сокращений разных мышц при сложных движениях, то есть координирует движения. Функции мозжечка наглядно иллюстрируются симптомами мозжечковых нарушений:
- астения – снижение силы мышечных сокращений, быстрая утомляемость;
- астазия – утрата способности к длительному тоническому сокращению мышц, что затрудняет сохранение позы;
- дистония – непроизвольное изменение мышечного тонуса;
- тремор – дрожание, усиливающееся при движении;
- дисметрия – расстройство размерности движений (избыточные или недостаточные движения);
- атаксия – нарушение координации движений, невозможность их выполнения в нужном порядке. Проявлениями атаксии являются адиадохокинез – затруднение смены направления вращательных движений, асинергия – нарушения содружественного сокращения мышц, в результате чего человек не способен, например, сесть из положения лежа без помощи рук, пьяная походка;
- дизартрия – нарушение речевой моторики.
При мозжечковых нарушениях главным образом страдают движения, которым человек обучался в течение жизни, следовательно, мозжечок участвует в формировании двигательной памяти.
Пирамидная и экстрапирамидная системы действуют совместно и согласованно. Пирамидная система в процессе эволюции сформировалась позже в связи с усложнением и совершенствованием двигательного поведения. Передние пирамидные пути, заканчивающиеся на мотонейронах кистевых мышц, имеются только у приматов. При поражении пирамидных путей экстрапирамидная система может частично компенсировать двигательные нарушения – произвольные движения, кроме самых тонких и точных, могут восстанавливаться.
Лекция 11. РЕГУЛЯЦИЯ ВИСЦЕРАЛЬНЫХ ФУНКЦИЙ.
ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ (АВТОНОМНОЙ) НЕРВНОЙ СИСТЕМЫ
Гуморальная и нервная регуляция. Эндокринные функции гипоталамуса
Разделение функций организма на анимальные (соматические) и вегетативные (висцеральные) было предложено французским анатомом и физиологом Ф.Биша в начале XIX века. Соматические функции – связь с внешнгей средой и движение – регулируются соматической нервной системой (СНС), включающей сенсорные и двигательные системы. Регуляция висцеральных функций происходит при сложном взаимодействии нервных и гуморальных механизмов. Гуморальная регуляция осуществляется разнообразными веществами, перемещающимися с кровью и тканевой жидкостью, нервная - вегетативной, или автономной, нервной системой (ВНС).
Гуморальные механизмы, в сравнении с нервными, более древние, их эффекты диффузны, развиваются медленнее и сохраняются дольше. Центром объединения нервного и гуморального контроля является гипоталамус. Многие ядра гипоталамуса способны к нейросекреции. Гипоталамус выделяет рилизинг-гормоны (либерины и статины), влияющие на функцию гипофиза и оказывающие самостоятельное влияние на ЦНС. Например, тиролиберин повышает уровень бодрствования, оказывает антидепрессивное действие, усиливает работу дыхательного центра. Соматостатин оказывает противоположное действие, снижает уровень эмоциональности и болевую чувствительность, оказывает тормозящее влияние на многие системы организма («всеобщий ингибитор»). Гонадолиберины оказывают стимулирующеевлияние на половое поведение, повышают эмоциональность, улучшают обучение. Кортиколиберин стимулирует двигательную активность и эмоциональность, но тормозит пищевое и половое поведение. В гипоталамусе синтезируются также нейропептиды, выделяющиеся в кровь через заднюю долю гипофиза и имеющие собственное влияние на висцеральные системы. Окситоцин вызывает сокращение гладких мышц органов репродуктивной системы при родах, оргазме, лактации, ухудшает обучение и память. Вазопрессин, или антидиуретический гормон (АДГ) – усиливает реабсорбцию воды в почечных канальцах, вызывает сужение кровеносных сосудов, в малых дозах улучшает память и обучение.
Вегетативная нервная система регулирует обмен веществ, рост и размножение (трофическая функция), координирует работу органов и систем (адаптационная функция). Адаптационно-трофическое влияние ВНС распространяется на все отделы НС, включая и кору головного мозга. Такая обратная связь превращает нервную систему в систему управления замкнутого цикла, а весь организм представляет собой саморегулирующуюся систему от клеточного до организменного уровня.
Все многообразные адаптационные реакции организма можно разделить на две группы – эрготропные и трофотропные, в процессе жизнедеятельности постоянно сменяющие друг друга и имеющие разную степень выраженности в зависимости от условий возникновения реакции.
|
|
|
