 |


Артериальные хеморецепторы
|
|
|
|
Периферические или артериальные хеморецепторы расположены в известной рефлексогенной зоне – дуге аорты и каротидном синусе (рисунки 17А и Б), и представлены каротидными и аортальными телами. Здесь же локализованы и барорецепторы, принимающие участие в регуляции артериального давления.
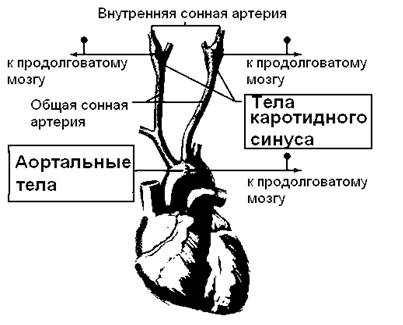
Рисунок 17 А. Периферические хеморецепторы
В сосудистой рефлексогенной зоне
Из двух хеморецептивных зон артериального русла - аортальной и синокаротидной - в регуляции дыхания существенную роль играет синокаротидная зона. Эта роль значительно скромнее по сравнению с ролью бульбарных структур - у человека двустороннее удаление каротидных тел не вызывает заметных изменений дыхания в состоянии покоя. Каротидные тела расположены в месте деления общей сонной артерии на внутреннюю и наружную.
Тело представляет собой образование, заключенное в соединительнотканную капсулу, чрезвычайно богато кровоснабжается и иннервируется как афферентными, так и эфферентными нервами. Кровоток через каротидное тело очень велик - до 2л/мин/г, а потребление кислорода в 3 - 4 раза больше, чем мозгом.

Рисунок 17 Синокаротидная (а) и аортальная (б) рефлексогенные зоны
IX и X – языкоглоточный и блуждающий нервы, 1 – верхний шейный симпатический ганглий, 2 – синусный нерв, 3 – каротидное тело, 4 – общая сонная артерия, 5 - затылочная артерия, 6 – звездчатый ганглий, 7 – аортальный нерв, 8 – аортальные тела, 9 – дуга аорты
Схема строения и иннервации каротидного тела представлена на рисунке 18.

Рисунок 18. Схема строения каротидного тела
1. Клетки I типа
2. Клетки II типа
3. Синусный нерв
4. Афферентные волокна синусного нерва
|
|
|
5. Эфферентное волокно синусного нерва
6. Симпатическое волокно
7. Кровеносный сосуд
В ткани каротидного тела различают два типа клеток. I тип - главные клетки, крупные клетки, имеющие эпителиальное происхождение. В клетках этого типа содержатся гранулы, которые исчезают при острой гипоксии. Непосредственно с ними контактируют окончания афферентной ветви языкоглоточного нерва (нерв Геринга, синусный нерв). Именно этим клеткам принадлежит основная роль в хемочувствительности - разрушение этих клеток прекращает хеморецептивную активность каротидного тела. Мелкие клетки II типа гомологичны глиальным клеткам и напоминают Шванновские. Своими отростками они оплетают главные клетки.
Адекватными стимуляторами хеморецепторов каротидного тела служат следующие сдвиги в составе омывающей их артериальной крови: 1) снижение напряжения кислорода, 2) увеличение напряжения СО2, 3) увеличение концентрации водородных ионов.
Главным стимулятором активности каротидного центра является гипоксия. Даже умеренная гипоксия сопровождается более выраженным увеличением частоты импульсов синусного нерва, чем сильная гиперкапния.
Каким же образом рецепторы воспринимают информацию о снижении напряжения кислорода в крови? Цитоплазма клеток I типа содержит гранулы, в которых накапливается дофамин. Оценка уровня кислорода осуществляется специальными рецепторами, которые расположены на мембране клеток I типа. На основании экспериментальных данных предложена гипотетическая схема работы этих рецепторов, представленная на рисунке 19.
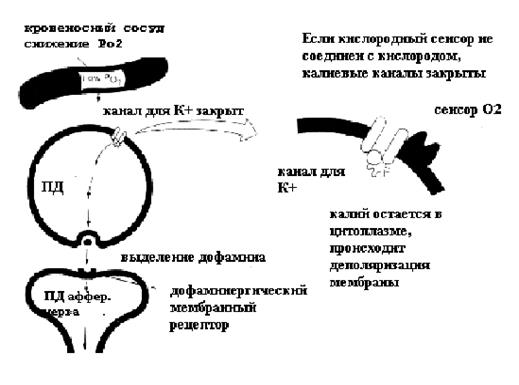
Рисунок 19. Кислородный сенсор каротидного тела
Взаимодействие кислородного сенсора с кислородом приводит к активации калиевых каналов. В таком состоянии клетка находится практически постоянно, а ток калия из цитоплазмы поддерживает потенциал клетки на уровне мембранного потенциала покоя. Снижение напряжения кислорода в крови приводит к освобождению кислородного сенсора, калиевые каналы закрываются, мембранный потенциал уменьшается и достигает критического уровня деполяризации, в клетках I типа возникает потенциал действия. Возникновение ПД приводит к тому, что в клетках открываются кальциевые каналы и выделяется дофамин.
|
|
|
Артериальные хеморецепторы возбуждаются и при повышении напряжения углекислого газа в артериальной крови. Гиперкапническая стимуляция артериальных хеморецепторов, так же как и центральных, осуществляется прямым влиянием ионов Н+ при снижении рН крови. Воздействие водородных ионов в клетках каротидного тела обусловлено сдвигом метаболизма за счет работы редокс-систем. Таким образом, и гипоксия, и гиперкапния различными путями приводят к изменению метаболических процессов в клетках, а стимуляторами каротидных хеморецепторов служат продукты измененного обмена. Существенная и важная разница заключается в том, что реакция на снижение напряжения кислорода наступает значительно быстрее.
Возникающий импульс возбуждения проводится по аффрентным волокнам синусного нерва и достигает дорзальной группы дыхательных нейронов продолговатого мозга. Возбуждение нейронов повышает инспираторную активность. Особенно увеличивается частота импульсации в диапазоне напряжения кислорода от 80 до 20 мм рт.ст.
Хеморецепторы каротидного синуса находятся под нервным контролем: повышение активности симпатической нервной системы и выделение норадреналина повышает их чувствительность, а парасимпатические импульсы и ацетилхолин – снижают.
Аортальные тела сходны по строению с каротидными телами, не отличаются и важнейшие функции этих образований, прежде всего как кислородных сенсоров. Расположенные в аортальной зоне хеморецепторы принимают незначительное участие в регуляции дыхания, их основная роль проявляется в регуляции деятельности сердца и тонуса сосудов.
Периферические хеморецепторы дополняют деятельность центральных. Взаимодействие центральных и периферических структур особенно важно в условиях дефицита кислорода. Дело в том, что центральные хеморецепторы очень чувствительны к недостатку кислорода. Клетки при гипоксии могут совсем потерять свою чувствительность, при этом снижается активность дыхательных нейронов. В этих условиях дыхательный центр получает основную возбуждающую стимуляцию от периферических хеморецепторов, для которых основным стимулом является именно дефицит кислорода. Таким образом, артериальные хеморецепторы служат «аварийным» механизмом стимуляции дыхательного центра в условиях снижения снабжения мозга кислородом.
|
|
|
Непременным условием эффективности легочного газообмена служит поддержание оптимальных вентиляционно-перфузионных отношений. Такое оптимальное соотношение обеспечивается сопряженной регуляцией систем дыхания и кровообращения. Проявлением такого сопряжения служит одновременное увеличение и вентиляции легких (МОД), и сосудистого тонуса, и деятельности сердца (МОК). Особенно выраженными такие одновременные изменения бывают во время физической нагрузки, при гипоксии, эмоциональном возбуждении. Периферические хеморецепторы расположены в тех же зонах, что и барорецепторы - нервные окончания лежащие непосредственно в стенке магистрального сосуда. Такое соседство, конечно, не случайно. Совместный контроль дыхания и кровообращения обеспечивает устойчивое снабжение кислородом жизненно важных органов, прежде всего мозга. Аортальная зона расположена у «ворот» всей артериальной системы, и здесь ведущую роль играют барорецепторы. Синокаротидная зона лежит у «ворот» всей сосудистой сети мозга, и здесь основная роль принадлежит хеморецепторам. Проекции хеморецепторных и барорецепторных афферентных волокон в ядрах ствола мозга (солитарное, парамедиальное) объединены межнейронными связями.
Итак, центральные и периферические хеморецепторы передают в дыхательный центр информацию о напряжении кислорода и углекислого газа в крови, они возбуждаются и увеличивают частоту импульсов при снижении содержания кислорода и повышении углекислого газа.
Механорецепторы
Дыхательный центр получает информацию и от механорецепторов. Эта информация поступает от чувствительных нервных окончаний, расположенных в гладких мышцах, в подслизистом слое и эпителии воздухоносных путей. Выделяют три основные вида рецепторов: 1) рецепторы растяжения легких, 2) ирритантные рецепторы, 3) J - рецепторы - юкстакапиллярные рецепторы легких
|
|
|
Рецепторы растяжения легких расположены в гладких мышцах воздухоносных путей - в трахее и бронхах всех калибров. В каждом легком около 100 таких рецепторов, они связаны с дыхательным центром толстыми миелинизированными волокнами блуждающего нерва с высокой (40м/сек) скоростью проведения. Возбуждение этих рецепторов возникает или усиливается при увеличении объема легких. Чем глубже вдох, тем больше частота импульсов, посылаемых рецепторами растяжения в дыхательный центр. Рецепторы обладают разными порогами возбуждения. Часть рецепторов обладает низким порогом - эти рецепторы возбуждены в течение выдоха и даже при спадении легких, во время вдоха частота импульсов резко возрастает. Высокопороговые рецепторы возбуждаются только при увеличении объема легких сверх функциональной остаточной емкости. Таким образом, информация от механорецепторов поступает в дыхательный центр на протяжении всего дыхательного цикла, этому способствует и медленная адаптация рецепторов.
Увеличение объема легких стимулирует рецепторы растяжения косвенно. Непосредственным раздражителем является внутреннее напряжение стенки воздухоносных путей. Это напряжение зависит от разности давлений по обе стороны стенки. При увеличении объема легких возрастает и их эластическая тяга. Альвеолы, стремящиеся к спадению, растягивают стенки бронхов в радиальном направлении. Поэтому возбуждение рецепторов зависит не только от объема легких, но и от свойств легочной ткани, ее растяжимости.
Большая часть афферентных волокон от рецепторов растяжения направляется в дорзальное дыхательное ядро продолговатого мозга, в этом ядре активность инспираторных нейронов изменяется не одинаково, около 60 % этих нейронов тормозится, тормозится и инспирация в целом. Таким образом, рецепторы растяжения легких обеспечивают обратную связь между дыхательным центром и глубиной дыхания (ДО). Рецепторы сигнализируют об объеме легких и скорости его изменения. Способность рецепторов растяжения возбуждаться при увеличении объема легких лежит в основе рефлекторной саморегуляции дыхания, в соответствии с которой длительность фаз дыхательного цикла и дыхательный объем определяются частотой импульсов от механорецепторов. Возбуждение рецепторов растяжения легких вызывает рефлекторное торможение вдоха и переход к выдоху. Этот рефлекс называется инспираторно-тормозящим рефлексом Геринга-Брейера. Следует отметить, что у взрослого человека рефлексы Геринга-Брейера включаются, когда дыхательный объем превышает 1 литр (например, при физической нагрузке), тормозной рефлекс проявляется так же во сне или под наркозом.
|
|
|
Ирритантные рецепторы - это быстро адаптирующиеся рецепторы воздухоносных путей, слизистой оболочки трахеи и бронхов. Особенно много их у корней легких. Ирритантные рецепторы обладают одновременно свойствами и механо- и хеморецепторов. Пороги возбуждения этих рецепторов выше, чем у рецепторов растяжения, они возбуждаются лишь при резких изменениях объема легких. Кроме того, ирритантные рецепторы возбуждаются при действии пылевых частиц, накапливающейся в воздухоносных путях слизи, при действии паров едких веществ (аммиака, эфира, табачного дыма), некоторых биологически активных веществ, например, гистамина. Это быстро адаптирующиеся рецепторы. При раздражении рецепторов этого типа в трахее - возникает кашель, неприятные ощущения типа першения, жжения, при возбуждении ирритантных рецепторов бронхов дыхание становится частым и поверхностным. Кроме того, импульсы от этих рецепторов вызывают рефлекторное сужение бронхов - бронхоконстрикцию.
Вблизи от капилляров малого круга кровообращения в интерстициальной ткани альвеол расположены J -рецепторы - юкстакапиллярные рецепторы легких. Раздражителем для этих рецепторов является повышение артериального давления в малом круге, увеличение объема интерстициальной жидкости в легких, эти рецепторы чувствительны к некоторым биологически активным веществам, например простагландинам, гистамину, никотину. Импульсы от этих рецепторов направляются к дыхательному центру по медленным немиелинизированным волокнам блуждающего нерва. Раздражение этих рецепторов приводит к частому, поверхностному дыханию и бронхоконстрикции. Сильное и устойчивое возбуждение J-рецепторов наблюдается при застое крови в малом круге, отеке легких, закупорке мелких сосудов и других повреждениях легочной ткани.
Информация от всех перечисленных рецепторов поступает в дыхательный центр, расположенный в продолговатом мозге.
Дыхательный центр
Дыхательный центр это совокупность связанных между собой нейронов центральной нервной системы, обеспечивающих 1) координированную ритмическую деятельность дыхательных мышц и 2) приспособление дыхания к меняющимся условиям окружающей и внутренней среды. Дыхательный центр был описан в 1885 году Н.А. Миславским. Этот центр, как и любой нервный центр, не является абсолютно автономным образованием, он представляет часть функциональной системы, регулирующей параметры внутренней среды - напряжение кислорода, углекислого газа и рН, и обеспечивает ее гомеостаз.
Мы уже упоминали о том, что в продолговатом мозге на дне IV желудочка расположены структуры, разрушение которых уколом иглы ведет к прекращению дыхания и гибели организма. В экспериментах с перерезками ствола мозга на различных уровнях было установлено, что центр, расположенный в продолговатом мозге является важнейшим в регуляции дыхания. Основной вклад в изучение центральных механизмов регуляции дыхания в последние годы вносят эксперименты с регистрацией активности отдельных нейронов. В ствол мозга вводят внутриклеточные или внеклеточные микроэлектроды, регистрируют активность отдельных нейронов и сопоставляют ее с одновременной записью дыхательных движений. Применение микроэлектродной техники позволило установить, что в состав дыхательного центра входят несколько типов клеток, активность которых соответствует фазам дыхательного цикла. Эти клетки были названы дыхательными нейронами.
Среди дыхательных нейронов выделяются две основные популяции клеток: к первой относятся нейроны, возбуждение которых совпадает с фазой вдоха - инспираторные нейроны, ко второй относятся нейроны, которые возбуждаются в фазу выдоха - экспираторные нейроны.
Природа ритмической активности дыхательного центра до настоящего времени не полностью понятна. Мы приведем гипотезы, существующие по этому вопросу, а в дальнейшем - факты, подтверждающие или опровергающие их.
1. Инспираторные нейроны спонтанно непрерывно активны и периодически тормозятся экспираторными. Экспираторные нейроны возбуждаются влияниями блуждающего нерва и вышележащих отделов мозга.
2. Обе группы нейронов спонтанно активны, а между ними существуют реципрокные отношения (это значит, что при возбуждении одной группы нейронов другая затормаживается, и наоборот).
3. Дыхательные нейроны не обладают спонтанной активностью, а возбуждаются от других отделов мозга. Обе группы нейронов связаны реципрокными отношениями через тормозные нейроны.
4. Существует нейронная сеть, содержащая несколько подтипов нейронов. Их взаимодействие приводит к ритмической активности всей сети. Возникновение этой ритмической активности обусловлено импульсами, поступающими от хеморецепторов и активирующей ретикулярной формации продолговатого мозга.
Как видим, существуют две противоположные точки зрения относительно спонтанной активности дыхательных нейронов.
|
|
|
