 |


Регуляции вентиляции легких по принципу отклонения
|
|
|
|
1. Дыхательный центр продолговатого мозга
2. Хеморецепторы каротидного синуса
3. Хеморецепторы продолговатого мозга
4. Рецепторы растяжения легких
5. Диафрагма
6. Межреберные мышцы
7. Увеличение размеров грудной клетки при сокращении диафрагмы и наружных межреберных мышц

Ритмический процесс чередования вдохов и выдохов определяется поэтапным включением нейронов, начиная от ранних инспираторных к поздним экспираторным. Этот циклический процесс у здорового человека в покое ограничивается только сокращением инспираторных мышц, следовательно, только возбуждением бульбоспинальных инспираторных нейронов, выдох протекает пассивно. Вместе с тем, в экспираторных нейронах во время выдоха всегда регистрируется активность в виде ВПСП, которые свидетельствуют об активности этих нейронов. Попытаемся, используя имеющуюся информацию о нейронах дыхательного центра усложнить схему и отобразить взаимодействие различных нейронов в организации дыхательного цикла.
Обсуждая взаимодействие дыхательных нейронов, попытаемся ответить на три важных вопроса: 1) какой механизм запускает инспирацию, 2) останавливает инспирацию и обеспечивает переход к расслаблению инспираторных мышц, 3) запускает активную экспирацию?
Механизм, который запускает инспирацию, назван центральной инспираторной активностью (ЦИА). Как уже было сказано, механизм ЦИА на сегодняшний день окончательно не понят, единственное, что установлено - отсутствие истинного водителя ритма в дыхательном центре. Казалось бы, инспирация запускается импульсами от периферических и центральных хеморецепторов при увеличении напряжения углекислого газа и (или) снижении напряжения кислорода в крови. В этом случае на роль ЦИА подошли бы ранние инспираторные нейроны, которые активируются при возбуждении хеморецепторов.
|
|
|
Однако, как уже упоминалось, в покое напряжение газов в периферической крови остается постоянным. Импульсы от хеморецепторов, конечно, поступают в дыхательный центр, но это поступление импульсов постоянное - тоническое, а включение вдоха происходит ритмически, следовательно, необходимо поискать еще какой-то стимул для возбуждения ранних инспираторных нейронов. Вспомним, что эти нейроны находятся в реципрокных отношениях с ранними экспираторными нейронами (тормозятся при их активации) и снижают свою активность при увеличении импульсов от рецепторов растяжения легких. Таким образом, стимулом может быть сочетание двух факторов: увеличение потока импульсов от хеморецепторов и снижение такого потока от рецепторов растяжения легких. Снижение эффекта рецепторов растяжения легких воздействует на ранние инспираторные нейроны двумя путями: и непосредственно затормаживая их, и возбуждая ранние экспираторные нейроны. Ранние инспираторные нейроны получают еще один тормозящий их поток импульсов - от поздних инспираторных нейронов. Если вспомнить о том, что П-И нейроны растормаживаются по мере нарастания инспирации и тоже находятся в реципрокных отношениях с ранними инспираторными, то можно говорить еще об одном стимулирующем влиянии на ранние: отсутствие активности поздних инспираторных нейронов. Активность этих нейронов нарастает по мере развития вдоха под возбуждающим влиянием инспираторных нейронов с нарастающей активностью. (ПН-И), которые возбуждаются от: 1) ранних инспираторных, 2) рецепторов растяжения легких, 3) периферических и центральных хеморецепторов. Вероятно, ранние инспираторные нейроны, суммируя на своих телах и дендритах возбуждающие и тормозные влияния от хемо- и механорецепторов, поздних инспираторных и ранних экспираторных нейронов, возбуждаются тем интенсивнее, чем выше стимуляция от хеморецепторов и ниже от рецепторов растяжения легких. Такое сочетание влияний можно рассматривать как ЦИА - центральную инспираторную активность, во всяком случае, до тех пор, пока не будет найдено новых экспериментальных данных, опровергающих это предположение. На схеме(рисунок 23) обозначим этот блок номером I, и будем считать, что в основе ЦИА лежит сочетание возбуждения от хеморецепторов, отсутствие возбуждения от рецепторов растяжения легких, торможение поздних инспираторных и ранних экспираторных нейронов - фактически это состояние выдоха, которое и стимулирует следующий вдох.
|
|
|
Рассмотрим блок под цифрой II на схеме. Активность нейронов с нарастающей активностью (ПН-И) поддерживается импульсами от: 1) ранних инспираторных, 2) рецепторов растяжения легких, 3) периферических и центральных хеморецепторов. Если вспомнить, что поздние инспираторные нейроны находятся с ранними в реципрокных отношениях, становится ясно, что интенсивность ЦИА определяет и интенсивность вдоха - его глубину, и его длительность. Глубина определяется интенсивностью возбуждения инспираторных нейронов с нарастающей активностью, а длительность - растормаживанием поздних инспираторных нейронов. Таким образом, чем глубже вдох (активность ЦИА), тем позже начнется прекращение инспирации, потому что поздние инспираторные нейроны дольше будут заторможены. Следовательно, второй блок на нашей схеме - это механизм, который определяет и время окончания инспирации, и соответствие между глубиной и частотой дыхания – чем глубже дыхание, тем оно реже.
Наконец, механизм активной экспирации (III). Пожалуй, мы его уже объяснили, подведем лишь итог: ранние экспираторные нейроны активированы во время вдоха, но их ВПСП не достигают критического уровня. Чтобы эти деполяризующие потенциалы перешли в потенциалы действия необходимо, как ясно из предыдущих рассуждений, 1) затормозить ранние инспираторные нейроны, 2) усилить поток импульсов от рецепторов растяжения легких. Ранние инспираторные нейроны затормаживаются при увеличении импульсации от рецепторов растяжения легких и при повышении активности поздних инспираторных. Таким образом, чем глубже вдох (активность ЦИА), тем интенсивней поток импульсов к ранним экспираторным нейронам от ранних инспираторных, тем больше вероятность формирования ПД на телах этих нейронов и активной экспирации.
|
|
|
Выдохи становятся активными при увеличении вентиляции легких, обусловленной различными причинами: усилением раздражения медуллярных и артериальных хеморецепторов, физической работой, при громкой речи, крике, при произвольной гипервентиляции и глубоких вдохах. Первая, постинспираторная фаза, в таких условиях укорачивается или исчезает. Активные выдохи обеспечивают усиление экспираторного тока воздуха. Их энергия суммируется с эластической энергией легких, накопленной во время вдоха.
Следовательно, на уровне нейронов дыхательного центра продолговатого мозга формируется паттерн дыхания, в котором учитываются и влияния рецепторов, и сила и длительность возбуждения отдельных групп нейронов. Поэтому, вероятно, задачей отдельных нейронов является и объединение в сеть для формирования последовательности фаз дыхательного цикла, и получение каждым типом нейронов своего потока информации и изменение паттерна дыхания в соответствии с этим потоком.

Рисунок 23. Схема организации дыхательного цикла
 Р-И- ранние инспираторные,
Р-И- ранние инспираторные,
ПН-И- полные инспираторные с нарастающей активностью,
П-И – поздние инспираторные,
Р-Э – ранние экспираторные,
П-Э –поздние экспираторные
проприобульбарные нейроны продолговатого мозга. Эти нейроны организуют паттерн дыхания.
Пневмотаксический центр
Структуры продолговатого мозга обладают способностью генерировать дыхательный ритм на основании информации от хемо- и механорецепторов. Однако после перерезки мозга ниже варолиевого моста у экспериментальных животных дыхательный ритм изменяется. У животных наблюдается длительный выдох, который редко прерывается резким вдохом, такое дыхание называется гаспинг. Следовательно, для полноценного дыхания необходимы структуры, расположенные в районе моста (рисунок 24). Структурам моста, необходимым для поддержания полноценного дыхания Люмсден в 1923 году дал название пневмотаксический центр (ПТЦ). Как уже было отмечено, пневмотаксический центр расположен в дорсолатеральной области ростральной части моста. Функции этого центра изучены с помощью физиологических экспериментов, включающих повреждение центра, его раздражение и отделение от продолговатого мозга путем перерезки. Повреждение центра сопровождается уменьшением вентиляции. Результат перерезки уже описан - длительный выдох, который редко прерывается вдохом - гаспинг. Интересно, что похожие результаты получены в экспериментах с одновременной перерезкой блуждающих нервов и повреждением структур моста. У таких животных наблюдается длительный вдох, изредка прерываемый коротким выдохом. Такой вариант дыхания называется апнейзис (рисунок 26 приложения). Эти результаты наводят на мысль о том, что ПТЦ и рецепторы растяжения легких оказывают однонаправленные влияния на дыхательный центр продолговатого мозга. Суть этих влияний заключается в облегчении переключения фаз дыхательного цикла. Это предположение подтверждается результатами экспериментов с раздражением ПТЦ. Раздражение пневмотаксического центра влияет на переключение фаз дыхания: при стимуляции ПТЦ инспираторная активность и усиливается, и раньше прекращается. Таким образом, в районе моста находится нейронный механизм, который стабилизирует деятельность дыхательного центра. Этот механизм, подобно импульсам от рецепторов растяжения легких, облегчает смену фаз дыхательного цикла.
|
|
|
Для того чтобы понять, каким образом ПТЦ принимает участие в регуляции дыхания, необходимо знать, откуда эти структуры получают информацию и куда направляют свои импульсы. К ПТЦ отправляются аксоны нейронов от дорсального и вентрального ядер продолговатого мозга. ПТЦ тоже посылает импульсы по аксонам нейронов в продолговатый мозг. Таким образом, между продолговатым мозгом и ПТЦ существуют двусторонние связи.
В ПТЦ тоже обнаружены дыхательные нейроны. Их немного и они организованы в небольшие группы по 10-12 нейронов. В отличие от дыхательных нейронов бульбарного центра, для которых характерна залповая активность, большинство нейронов ПТЦ обладает непрерывной (тонической) импульсной активностью. Частота импульсов совпадает с ритмом дыхательного цикла. Особенно много в ПТЦ нейронов, частота импульсов у которых резко увеличивается во время одной из фаз дыхательного цикла и уменьшается во время другой (инспираторно-экспираторные, экспираторно-инспираторные нейроны). Наибольшая частота разрядов фиксируется во время смены фаз. Такие нейроны получили название фазовоохватывающих, им приписывается функция связывания отдельных фаз дыхательного цикла. Важной особенностью нейронов ПТЦ является то, что при нарушении связей с бульбарным центром они теряют свою активность. Итак, первая группа тонических нейронов получает информацию от бульбарного центра. Это информация об активности различных групп дыхательных нейронов, которая в свою очередь определяется импульсами от хеморецепторов и рецепторов растяжения легких. На нейронах ПТЦ конвергируют импульсы от вышележащих отделов мозга, от нейронов ретикулярной формации. Импульсы от ПТЦ поступают к дыхательным нейронам продолговатого мозга, причем преимущественно к бульбоспинальным нейронам. В настоящее время точно не известно инспираторные или экспираторные нейроны находятся под влиянием ПТЦ, однако это не имеет существенного значения, если вспомнить о том, что деятельность генератора ритма организована по принципу циклического возбуждения. Следовательно, активация любой группы нейронов приведет к ускорению этой циклической деятельности. Эти предположения вполне соответствуют роли ПТЦ, определенной экспериментально - стабилизация и ускорение ритма, и облегчение переключения дыхательных фаз. Нейроны ПТЦ не участвуют в организации ритма дыхания, но способны изменить этот ритм на основании информации от самого дыхательного центра, от вышележащих отделов мозга и от ретикулярной формации ствола мозга.
|
|
|
В ПТЦ найдены и группы нейронов с фазной (импульсной) активностью. Оказалось, что фазная активность таких нейронов усиливается при переходе от сна к бодрствованию и при отсутствии раздражения от рецепторов растяжения легких. Предполагается, что роль таких фазных нейронов может заключаться в поддержании тонуса нейронов дыхательного центра в случае, если почему-то прекращается поток импульсов от рецепторов растяжения легких.
Таким образом, можно считать установленным, что между пневмотаксическим центром моста и дыхательным центром продолговатого мозга существует петля обратной связи. ПТЦ получает возбуждающие импульсы от дыхательных нейронов продолговатого мозга и, в свою очередь, активирует эти нейроны, стабилизирует деятельность дыхательного центра и облегчает переключение с одной фазы дыхательного цикла на другую. ПТЦ не является генератором дыхательной периодики.
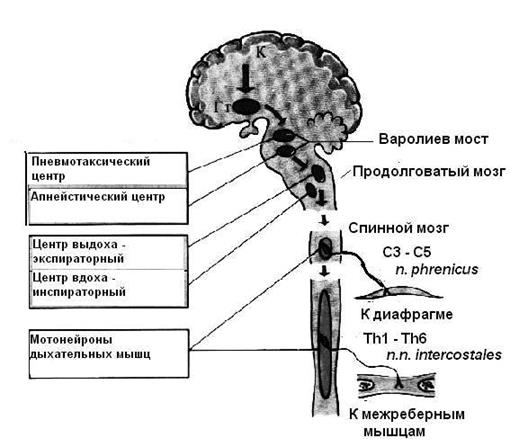
Рисунок 24. Локализация пневмотаксического
и апнейстического центров
В районе средней и каудальной частей моста существует еще один источник тонических влияний на дыхательный центр. Влияния этого центра противоположны влияниям ПТЦ. Этот центр назван апнейстическим центром и известно о его регулирующем влиянии на дыхание пока очень немного.
|
|
|
