 |


Морфофункциональная организация
|
|
|
|
Спинной мозг, являясь самым каудальным отделом центральной нервной системы, осуществляет две основные функции — рефлекторную и проводниковую. Импульсы, приходящие от экстерорецепторов кожи, проприо- и висцерорецепторов, обеспечивают разнообразие двигательных и вегетативных рефлекторных реакций, в характере протекания которых не последнюю роль играют межнейронные отношения в спинном мозгу и межцентральные отношения спинного и головного мозга, что обеспечивается внутриспиналь-ными межнейронными связями и восходящими и нисходящими проводящими путями спинного мозга.
Различные представители позвоночных обладают спинным мозгом неодинаковой длины. Так, у большинства рыб, хвостатых амфибий, рептилий и птиц он заполняет весь позвоночный канал, в то время как у бесхвостых амфибий и млекопитающих -- только часть этого канала. Самый короткий спинной мозг обнаружен у низших животных. Так, например, у южноамериканской жабы (Р1ра) конечный конус спинного мозга находится на уровне третьего позвонка, а у высокодифференцированной костистой рыбы (Мо1а) спинной мозг вообще не выходит за пределы черепа, и весь позвоночный канал заполнен лишь нервными волокнами.
Спинной мозг имеет сегментарное строение. От каждого сегмента отходят две пары корешков — вентраль-
ные и дорсальные, которые, соединяясь, образуют периферические спинномозговые нервы. В опытах с перерезками спинальных корешков, раздражением их и отведением от них электрической активности было показано, что задние корешки являются афферентными, чувствительными, центростремительными, а передние — эффекторными, двигательными (либо секреторными), центробежными -- это закон распределения корешков Белла—Мажанди. Перерезка задних корешков приводит не только к потере чувствительности, но и к нарушениям органов движения, которые становятся плохо координированными -- атаксическими. Спинномозговая атаксия возникает и у человека при поражениях восходящих путей спинного мозга. Такие расстройства координации движений наступают вследствие неправильного распределения мышечного тонуса в результате потери обратной связи от проприоре-цепторов и экстерорецепторов кожи во время осуществления двигательного акта.
|
|
|
Шеррингтоном в опытах с перерезками спинальных корешков было показано, что каждый сегмент спинного мозга иннервирует три метамера тела и получает афферентацию от трех метамеров, а каждый метамер иннервируется из трех сегментов спинного мозга и передает сигналы в три спинномозговых сегмента.
Соответственно распределению и функциям корешков на поперечном сечении спинного мозга различаются задние и передние рога центрального серого вещества, имеющего Н-образную форму. В задние рога входят волокна задних корешков (тела нейронов которых локализованы в спинальных межпозвоночных ганглиях), здесь распределены тела промежуточных нейронов (интернейронов) и находятся клетки жела-тинозной субстанции Роландо; в передних рогах ло-

|
| Кортикоспинальный тракт |
| Руброспияальный тракт |
| Латеральный р цти к улосп ин ал ьн ый тракт |
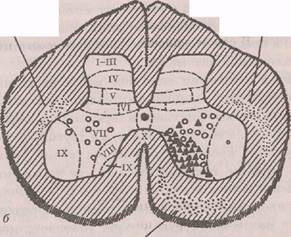
кализованы двигательные нейроны (мотонейроны), волокна которых покидают спинной мозг в составе передних корешков. Между дорсальными и вентральными рогами лежит промежуточная зона. Нервные клетки распределены и диффузно, и ядерно: медиальные и латеральные ядра — в переднем роге, ин-термедиомедиальное и интермедиолатеральное ядра — в промежуточной зоне, кларков столб и собственное ядро — в заднем роге. Интернейроны имеют небольшие размеры сомы, короткие дендриты и аксоны различной длины. Мотонейроны обладают крупной сомой, длинными ветвящимися дендритами, проникающими за пределы серого вещества, и длинными толстыми аксонами, иннервирующими мышцы. В боковых рогах спинного мозга локализованы вегетативные нейроны, представляющие собой инстанцию исполнительного механизма в регуляции вегетативных функций. В сером веществе спинного мозга интер- и мотонейроны образуют функциональные объединения, отвечающие за определенные специфические функции,— это функциональные единицы центральной нервной системы. Рекседом предложено разделение на XI пластинок (или слоев) серого вещества спинного мозга по анатомо-функциональным особенностям составляющих их нейронных элементов (рис. 4.1).
|
|
|
1-ГУ пластинки, образующие «головку дорсального рога», являются первичной сенсорной областью серого вещества для проекции большей части экстеро-цептивных афферентных волокон от туловища и конечностей. Из этой области идут тракты, передающие экстероцептивные сигналы к высшим центрам.
У-У1 пластинки образуют «шейку дорсального рога», где оканчиваются волокна от сенсомоторных областей коры и волокна проприоцептивной чувствительности туловища и конечностей; кроме того, в
Вестибулоспинальный тракт
 Латеральный ретикулоспиналыгый тракт (
Латеральный ретикулоспиналыгый тракт (
Медиальный ретикулоспиналышй тракт
Рис. 4.1. Схема разделения серого вещества спинного мозга у поясничного сегмента кошки (Рексед, 1964): а — расположение и области окончаний: слева — руброспинального тракта (светлые кружки); справа — вестибулоспинального (квадраты) а кортикоспинального трактов (треугольники и черные кружки); б — расположение окончаний медуллярного (светлые кружки) и мостового (треугольники) ретикулоспаналь-ных трактов (Шмидт, Тевс, 1983)
медиальную часть V (и IV) пластинки проецируются нисходящие волокна от сенсорных областей коры. По предположению Рекседа, здесь интегрируются рефлексы, обеспечивающие точную регуляцию движений.
Пластинка VII является областью обширных про-прио- и висцероспинальных связей, а также афферентных и эфферентных связей спинного мозга с мозжечком и средним мозгом. Часть VII пластинки, локализованная в вентральном роге, является тормозной областью, где преобладают клетки Реншоу.
|
|
|
Нейроны пластинки VIII, характеризующейся буль-бо- и проприоспинальными (особенно контралатераль-ными и комиссуральными) связями, могут модулировать моторную активность.
Пластинка IX является первичной моторной областью серого вещества и содержит мотонейроны спинного мозга, объединенные в функциональные группы (пулы) и представляющие собой общий конечный путь рефлексов спинного мозга.
Клетки нейроглии спинного мозга на значительном протяжении покрывают поверхность нейронов, причем отростки глиальной клетки направлены, с одной стороны, к телам нейронов, а с другой, часто контактируют с кровеносными капиллярами, являясь, таким образом, посредниками между нервными элементами и источниками их питания (Г. И. Поляков).
Спинной мозг по восходящим путям передает сигналы в надсегментарные уровни головного мозга, а но нисходящим получает оттуда команды к действию. Восходящие пути передают импульсы от проприоцеп-торов по волокнам спинобульбарных пучков Голля и Бурдаха (задние столбы белого вещества) и спиномоз-жечковых путей Говерса и Флексига (боковые столбы), от болевых и температурных рецепторов по латеральному спиноталамическому тракту, от тактильных
рецепторов по вентральному спиноталамическому пути и частично по пучкам Голля и Бурдаха.
Нисходящие пути проходят в составе кортикоспи-нальных, или пирамидных, трактов (передние и боковые столбы) и экстракортикоспинальных, или экстрапирамидных: ретикулоспинального, руброспи-нального, тектоспинального, вестибулоспинального, оливоспинального пучков.
4.2. Рефлекторная деятельность спинного мозга
Характер рефлекторной реакции спинного мозга и скрытый период рефлекса зависят от ряда условий: от модальности возбуждаемых рецепторов, от обширности возбуждаемой рефлексогенной зоны, от скорости проведения в нервных волокнах, от интенсивности и продолжительности раздражения, а также от функционального состояния нервной системы.
|
|
|
Полная поперечная перерезка спинного мозга вызывает спинальный шок, выражающийся в полном выпадении рефлекторных функций отделов, локализованных ниже места перерезки. В пользу того, что причиной шока является отторжение от головного мозга нижних этажей спинного, говорит отсутствие нового шока при повторной перерезке спинного мозга (ниже места прежней перерезки), которая производится после частичного восстановления функций в этой части мозга (Шеррингтон).
Глубина и длительность спинального шока значительно более выражены у животных с существенным доминированием энцефализации. Так, у лягушек спинальный шок длится всего несколько минут, у хищных млекопитающих — несколько часов, у обезьян — несколько дней или недель, у антропоидных обезьян и человека — несколько месяцев.
Есть предположение, что перерыв нисходящих путей спинного мозга приводит к подавлению рефлек-
сов за счет выключения большого количества возбуждающих влияний на а-, р-мотонейроны и другие спи-нальные нейроны, а также за счет растормаживания тормозных связей. Возникающая через определенное время после шока компенсация спинальных функций может привести к гиперрефлексии, которая является следствием выпадения тормозящих влияний головного мозга, в частности, его ретикулярной формации. Однако все же механизмы спинального шока, а тем более постшокового восстановления функций до конца еще не ясны.
Рефлекторная деятельность спинного мозга часто протекает на фоне иррадиации возбуждения (Пфлю-гер, Шеррингтон), идущей и вдоль спинного мозга (в краниальном направлении на более дальние сегменты, чем в каудальном), и внутри сегмента на противоположную сторону. Характер и степень иррадиации зависят как от локализации и силы раздражения, так и от возбудимости центрального аппарата спинного мозга. В этом процессе принимают участие и промежуточные нейроны, активирующие в обычных условиях, в основном мотонейроны соответствующих сегментов, что и обеспечивает более или менее ограниченную рефлекторную реакцию при действии умеренного раздражения рецептивного поля. При повышении же возбудимости какого-либо координационного рефлекторного аппарата иррадиация возбуждения вызывает преимущественную активацию именно этого аппарата; такой процесс лежит в основе принципа доминанты (А. А. Ухтомский).
Одной из характерных черт рефлекторной деятельности спинного мозга (равно как и других отделов центральной нервной системы) является усиление рефлекторных реакций при повторных раздражениях (суммация), которое может рассматриваться как
|
|
|
результат повышения возбудимости в работающих нейронных системах. Повышение возбудимости центральных элементов рефлекторной дуги после одного залпа афферентных импульсов длится от нескольких сотен миллисекунд до нескольких секунд. Однако после тетанизирующего раздражения оптимальной частоты чувствительных волокон оно может длиться гораздо дольше {в течение минут) и выявляться тестирующим раздражением, дающим более высокий суммационный электрический ответ. Это — посттета-ническая потенциация (Ллойд). При иных же частотах тетанизирующего воздействия возбудимость рефлекторных элементов может снижаться, что приводит к посттетанической депрессии.
Электрическая активность
Активное состояние спинномозговых нейронов сопровождается изменением их электрогенеза. Это колебания двух типов: быстрые пиковые потенциалы продолжительностью в несколько миллисекунд, отражающие распространяющееся возбуждение в аксоне и соме, и медленные, растянутые на десятки миллисекунд, соответствующие локальным (синаптическим) процессам в соме и дендритах. При отведении же электрических колебаний от спинальной поверхности, спинномозговых корешков или из глубины спинного мозга регистрируются суммарные электрические эффекты, отражающие деятельность многих различных нервных элементов. Так, отводимая от переднего корешка активность представлена потенциалами действия аксонов, проходящих в составе корешка, и распространяющимися электротонически потенциалами сомы мотонейронов. От дорсальных корешков отводятся потенциалы действия заднекорешковых волокон и медленные потенциалы центральных окончаний афферент-

|
ных волокон, распространяющиеся сюда электрото-нически (Д. С, Воронцов). Отведение электрической активности от дорсальной поверхности спинного мозга или из глубины его заднего рога также обнаруживает сложную картину сочетания быстрых и медленных потенциалов. Отрицательное колебание (К-волна) сменяется длительным положительным колебанием (Р-вол-на), которое может рассматриваться как отражение длительной деполяризации центральных терминалей афферентных волокон (Экклс).
с
|
|
|
