 |


Значение модельных объектов в генетике поведения и психогенетике
|
|
|
|
Психогенетические исследования на современном этапе не ограничиваются изучением наследования тех или иных психофизиологических характеристик нервной системы животных и человека. Развитие молекулярно-генетических методов позволило выявить высокую степень гомологии (сходства, обусловленного общностью происхождения) многих генов человека и других млекопитающих. Гены, кодирующие жизненно важные функции, сохранялись почти неизменными (например, гены гемоглобинов, цитохромов и др.) на протяжении миллионов лет эволюции. Поэтому даже у эволюционно очень далеко отстоящих друг от друга организмов выявлена высокая степень гомологии между такими генами. В свою очередь это определяет гомологию некоторых организменных признаков, в том числе и поведенческих. Изучая работу генов, гомологичных человеческим, у более просто организованных организмов (у одноклеточных, нематоды, дрозофилы, мыши, крысы, собаки, обезьяны), можно успешно решать некоторые проблемы психогенетики. Более простое строение нервной системы в сочетании с широкими возможностями экспериментирования, включающими молекулярно-генетический и гибридологический анализ, клонирование и селекцию по заданным признакам, позволяют успешно изучать роль генов в процессах рецепции внешних воздействий, в контроле тонких механизмов формирования и проведения нервных импульсов, запоминания, научения и п.д.
Так, например, одним из модельных объектов, на котором изучают консерватизм генов, контролирующих общий план строения нервной системы, является Ciona intestinalis (хордовые беспозвоночные оболочники – асцидии). Тело личинки этой асцидии состоит из примерно 2600, а ее центральная нервная система - из 330 клеток. При этом более 230 клеток являются клетками глии, и менее 100 оставшихся - собственно нейронами. Такая простота организации нервной системы сравнима с данными для другого модельного объекта – нематоды Caenorhabditis elegans, у которой всего 302 нейрона. Создание коллекций мутантов по генам всех 6 хромосом C. elegans позволило многое понять в механизмах генетического контроля пищевого поведения, хеморецепции, локомоции и нейрогенеза (Корочкин, Михайлов, 2000; Corbo et al., 2001). Сравнительно-генетический анализ позволяет обнаружить гомологичные гены, контролирующие поведение, у нематоды, дрозофилы, мыши и человека (Salzberg, Bellen, 1996). Это может означать не только гомологию генных продуктов, но и их роли в процессах нейрогенеза и формировании девиантных форм поведения животных и человека.
|
|
|
Использование еще одного достаточно простого модельного объекта – морского брюхоногого моллюска – Aplysia (или морского зайца) в сопоставлении с данными на традиционных модельных объектах (мышь, дрозофила) позволяет глубже понять молекулярные механизмы формирования памяти и процесса научения. Несмотря на разбросанно-узловой тип нервной системы всего лишь из нескольких ганглиев, этот брюхоногий моллюск поддается простейшим формам обучения. Так, выработка условного рефлекса активного избегания прикосновений у Aplysia показала, что механизм передачи внешнего сигнала внутрь организма осуществляется, как и у высших организмов, путем активации в клетках эволюционно древнего и консервативного каскада биохимических реакций сигнальной трансдукции. В эти реакции вовлечены множество белков, гомологичных таковым у других животных, включая человека. В процессе реагирования на условный раздражитель у моллюска меняется сначала активность цитоплазматических ферментов, а затем, соответственно, генов раннего и позднего ответа. Этим определяются процессы возбуждения нервных волокон и проведения по ним электрических импульсов. Длительное «обучение» приводит к запоминанию: животное отвечает на прикосновение в несколько раз быстрее и помнит об этом длительное время. При этом меняется архитектоника нервных связей: у обученного моллюска от зоны раздражения по всей рефлекторной дуге образуются дополнительные синапсы. У морского зайца выявлено существование краткосрочной, среднесрочной и долгосрочной памяти. Анализ поведенческих отклонений (и коррелирующих с ними других эффектов) различных мутаций по гомологичным генам приближает нас к решению проблемы таких заболеваний, как болезнь Альцгеймера, рассеянность и старческое слабоумие.
|
|
|
Излюбленным объектом генетики является плодовая мушка – Drosophila melanogaster. Примерное число нервных клеток у дрозофилы составляет 250 тысяч: это значительно меньше, чем у мыши и человека (40 миллионов и 85 биллионов, соответственно). Коллекции мутантных форм дрозофилы по различным поведенческим признакам позволяют проводить их всесторонний генетический анализ. Среди единичных генов дрозофилы, которые затрагивают процессы обучения и запоминания можно, для примера назвать linotte, dunce («тупица»), rutabaga, amnesiac («без памяти»), radish, dCREB2-b и CXM. Примерами генов, влияющих на нейрогенез, являются: achaete, scute, Notch, Delta, cut, Prospero и многие др. Следует отметить, что для большинства из них уже найдены гомологичные гены у других животных и человека. Консервативность этих генов указывает на огромную адаптивную значимость нормальной работы нервной системы животных. Спонтанно возникающие мутации в таких генах, немедленно уничтожались в процессе естественного отбора как вредные, т. е. существенно снижающие приспособленность.
Гомология генов и определяемых ими признаков позволила успешно моделировать на дрозофиле некоторые нейродегенеративные заболевания. Целый ряд специфических мутаций по своему проявлению напоминает заболевания человека. Так, например, мутация drop-dead в гене drd приводит к преждевременному старению и смерти мух сходному с синдромами Вернера и Хатчинсона-Гилфорда у человека. Мутация «swiss cheese»в гене sws приводит к вакуолизации тканей мозга мух. Гомологичный генный продукт у позвоночных определяет развитие определенных нейропатий. Гомологом мутантной формы гена человека, контролирующего губчатую энцефалопатию, является мутация «sponge cake»дрозофилы. Найдены мутации, которые моделируют такие наследственные заболевания человека как болезнь Тея-Сакса, адренолейкодистрофия, болезнь Альцгеймера, семейный амиотрофический латеральный склероз и др. Использование современных методов по внедрению чужеродных генов в геном дрозофилы, позволило создать модели для изучения болезней Гентингтона и Мачадо-Джозефа.
|
|
|
Существенным преимуществом модельных объектов является их относительно короткий жизненный цикл. Некоторые нейродегенеративные заболевания развиваются у человека очень поздно (возрастные заболевания). Поэтому их трудно изучать и еще труднее выявить в раннем онтогенезе, чтобы начать своевременное лечение. Использование адекватных моделей на животных позволяет эффективно изучать механизмы развития таких заболеваний. Сходство биохимических механизмов развития подобных недугов приводит к тому, что некоторые лекарственные вещества, оказывают такое же действие на дрозофилу и мышь как на человека (или даже полностью вылечивают ее).
Чем ближе к человеку по уровню развития стоит лабораторный объект, тем ценнее получаемые исследователями данные по моделированию различных наследуемых форм поведения и их отклонений. Пионерами исследований в области научения животных были И.П. Павлов и американский физиолог Л. Торндайк. С момента первых работ Торндайка (1898) по обучению методом проб и ошибок у кошек и собак прошло всего около 10 лет, а в США уже было создано 8 зоопсихологических лабораторий, где проводили сравнительные исследования поведения на крысах, мышах, приматах и даже медузах и дождевых червях. С переоткрытием в 1900-е годы законов Г. Менделя стала бурно развиваться генетика. И хотя еще Ф. Гальтон был убежден, что интеллект и способности к обучению наследуются, только с 1920-х годов начинаются исследования по генетике поведения. Э. Толменом, а затем его учеником Р. Трайоном была показана возможность выведения (селекции) «умных» и «тупых» инбредныхлиний крыс, которые отличались друг от друга по скорости выработки условного рефлекса или обучения в лабиринте. Позднее аналогичные данные были получены на мышах и на других видах животных (например, на хомячках, гвинейских свинках, кроликах).
|
|
|
Примером могут служить линии мышей DBA/2J, BALB/c и C3H/He. Исследователями было показано, что при непрерывном обучении первая линия хорошо, а последняя плохо обучаются. Животные генотипа BALB/c демонстрировали в этих условиях промежуточный уровень правильных ответов. При «прерывистом» обучении, когда паузы между циклами обучения составляли от нескольких минут до 24 часов динамика процесса обучения у этих линий существенно различалась. Животные генотипа DBA/2J обучались успешнее, в то время как особи линии C3H/He уже при 2-часовом интервале полностью теряли способность обучаться. В тоже время на генотип BALB/c изменение схемы тренировок существенно не сказывалось. Анализ данных (Рис. 22) позволяет предполагать, что у животных генотипа C3H/He большая скорость кратковременного запоминания, но ослаблен процесс консолидации памяти. В тоже время у мышей линии DBA/2J долговременная память формируется достаточно быстро. Определенные схемы скрещиваний позволяют создавать, например, конгеннные линии лабораторных мышей. Использование таких линий позволило установить связь межу активнооборонительной реакцией с локусами гистонесовместимости Н-25 и W-56. Такая связь может указывать на то, что гены, контролирующие данную форму поведения, расположены вблизи этих районов.
Определения:
Инбридинг (у животных)– система близкородственных скрещиваний, ставящая целью получение особей, гомозиготных по всем генам (или, по-крайней мере, по изучаемым генам).
Инбредная линия – группа животных, полученная и поддерживаемая путем близкородственных скрещиваний на протяжении ряда поколений (для мышей и крыс – более 20 поколений братско-сестринского инбридинга). Считается (с определенной степенью допуска), что животные инбредной линии имеют идентичный генотип и гомозиготны по большинству генов.
Конгенные линии – линии, генотипы которых идентичны, за исключением аллелей одного гена (или группы тесно сцепленных генов) какого-либо одного небольшого района (локуса) хромосомы. Чаще всего термин конгенный применяется в отношении генов локуса гистонесовместимости, т.е. района сосредоточения генов, отвечающих за тканевую совместимость.
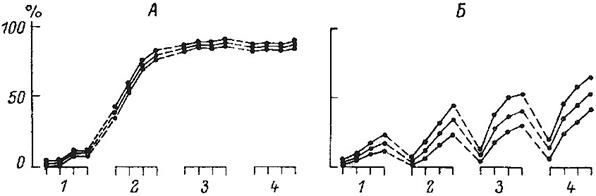 Рис 22. Динамика выработки условного рефлекса активного избегания у животных линий DBA/2J (А) и C3H/He (Б) в течение 24 дней с 24-часовыми интервалами между циклами обучения (Bovet et al., 1969)
По оси абсцисс – дни тестирования; по оси ординат – частота правильных ответов в процентах.
Рис 22. Динамика выработки условного рефлекса активного избегания у животных линий DBA/2J (А) и C3H/He (Б) в течение 24 дней с 24-часовыми интервалами между циклами обучения (Bovet et al., 1969)
По оси абсцисс – дни тестирования; по оси ординат – частота правильных ответов в процентах.
|
|
|
|
Сравнение некоторых форм поведения в созданных ранее инбредных линиях, отбиравшихся по медицинским и другим параметрам, также показало, что животные разных генотипов ведут себя по-разному. Было выявлено влияние генотипа на такие поведенческие характеристики животных как «эмоциональность», «исследовательское поведение», «агрессивность», «склонность к потреблению алкоголя», «двигательная активность», «способность к экстраполяции» и многие другие.
Так, например, линии лабораторных мышей существенно различаются по своему «отношению» к алкоголю (Табл. 5). В условиях выбора воды или раствора этанола средний уровень потребления алкоголя у животных одного генотипа может более чем в 5 раз превышать таковой у особей другой линии. При этом существенно отличается и тенденция к предпочтению или избеганию спиртового раствора в 3-х недельном эксперименте. В то время как в линиях С57BL/6 и С3Н/2 наблюдается постепенное увеличение потребления этанола, животные генотипов BALB/c и А/3 начинают все больше и больше предпочитать воду.
Развитие экспериментальных генетических и других методов позволило использовать химерных животных в качестве модели для изучения поведения. Так, С. Бензер, используя свойство кольцевой (аномальной) Х-хромосомы у плодовой мушки (Drosophila melanogaster), получал мух-мозаиков. Тело таких мух состоит из участков с двумя или только с одной Х-хромосомой. В нормальную Х-хромосому обычно вводят рецессивную аллель гена, отвечающего за какой-либо поведенческий (или другой) признак. Имея коллекции мозаичных особей со случайно комбинируемыми нормальными (ХХ) и мутантными (Х) участками тела можно изучать влияние мутантной аллели на определенные структуры в теле мозаика, которые определяют поведенческий или иной признак.
Табл. 5. «Предпочтение-избегание» потребления этанола в разных инбредных линиях лабораторных мышей
| Линия | Неделя | Спирт/Вода | Среднее (%) |
| C57BL | 0.085 0.093 0.104 | 9.4 | |
| C3H/2 | 0.065 0.066 0.075 | 6.9 | |
| BALB/C | 0.024 0.019 0.018 | 2.0 | |
| A/3 | 0.021 0.016 0.015 | 1.7 |
Некоторые мутации, в том числе затрагивающие функционирование нервной системы животных, летальны. Механизмы их действия трудно изучать традиционными методами. Энн Мак-Ларен была одним из первых исследователей, получившим химерных лабораторных мышей. Смешивая бластомеры разных эмбрионов она получала особей, части тела которых происходили от разных родительских пар. Создание жизнеспособных химер, у которых часть тела, состоящая из нормальных клеток, спасает весь организм от гибели, иногда может быть выходом из подобной ситуации: изучая, что именно (какой генный продукт) спасает химерную особь от гибели, можно понять механизм действия мутантного гена. Исследования показали, что при лечении болезни Паркинсона пациентам помогает подсадка в мозг эмбриональных дофаминэргических нейронов. Однако образование глиального рубца, изолирующего трансплантат, снижает успех лечения. Если же вместе с этими нейронами подсаживали эмбриональные клетки дрозофилы, образование глиального рубца задерживалось, и эффективность лечения возрастала. Таким образом, «химерная ткань» помогает мозгу пациента функционировать нормально.
Развитие методов молекулярной биологии и генной инженерии позволило создать принципиально новые подходы к изучению поведенческих признаков. С одной стороны, зная тонкое строение гена можно синтезировать его заведомо «порченные» копии, а затем замещать ими нормальные аллели в клетках будущего организма (генный нокаут). Затем остается только наблюдать за проявлениями такой мутантной аллели.
Можно создавать разные варианты одного и того же гена и сопоставлять их с отклонениями в проявлении признака. Самое интересное, что можно, оставляя сам ген неиспорченным, манипулировать регуляторными участками перед геном и позади него. Вставляя тканеспецифичные и отвечающие на какое-либо синтетическое вещество промоторы, можно создать аллель, которая будет работать только в определенной ткани и только по желанию экспериментатора. Ген будет включаться и выключаться при наличии в организме специфического вещества, которое появляется и исчезает под контролем исследователя, проводящего эксперимент («условный нокаут»). В мозге высших животных огромное число уникальных (т.е., тканеспецифичных) белков. Соответственно, работа таких генов регулируется тканеспецифичными промоторами. Создавая животных, «условно нокаутных» по генам, работающим в разных отделах мозга, можно изучать различные аспекты генетического контроля формирования их поведения. Так, например, были созданы мыши у которых только в поле СА1 гиппокампа был дефектен NMDA- рецептор глутамата - одного из главных нейромедиаторов мозга млекопитающих. Этого удалось добиться путем выключения в гиппокампе одного из генов, который кодирует субъединицу NR1 NMDA-рецептора. Такая модель позволила прояснить механизм формирования одного из видов нарушений памяти и обучения у животных и человека.
Интересную модель для психогенетических исследований представляют животные, находящиеся в стадии «одомашнивания». Процесс доместикации фактически представляет собой жесткий отбор животных в первую очередь по поведению, которое должно соответствовать желаниям человека. Эксперименты академика Д.К. Беляева и его коллег по одомашниванию лисиц показал, что отбор на неагрессивное по отношению к человеку поведение приводит к целому ряду гормональных, поведенческих и видимых простым глазом морфологических изменений. Некоторые из них идентичны признакам, наблюдаемым у собак, которые уже прошли длительный процесс «притирки» к человеку. Анализ изменений, происходящих во время одомашнивания различных видов, помог получить данные о тонких механизмах регуляции стрессоустойчивости (чувствительности) животных и человека.
Задания для самоконтроля:
1. Раскройте содержание понятия «гомологии». Приведите примеры гомологии генов и кодируемых ими признаков.
2. В чем преимущества использования модельных объектов в психогенетических исследованиях? Приведите примеры.
3. Расскажите об ограничениях в использовании модельных объектов. Приведите примеры.
4. Дайте определения инбридинга, инбредной и конгенной линии животных. Приведите конкретные примеры использования тех или иных линейных животных в психогенетических исследованиях.
5. Что такое память и научение? Каковы основные результаты применения современных молекулярно-генетических методик при изучении механизмов памяти?
6. Как в генетике поведения были использованы «мозаики» и «химеры»?
7. Какие современные молекулярно-генетические методы существенно расширяют возможности психогенетических исследований?
|
|
|
